Abstract
Patagonia is a region that has fascinated researchers for centuries considering the evidence of early human occupation, its geographical and environmental variability, and the diversity of human adaptations. From an archaeological and bioanthropological perspective, the region has been the focus of many studies addressing a wide range of questions, from a broad scale, such as the peopling of the Americas, to a local scale concerning the diversity and interactions of human populations. For three decades, paleogenetic studies have contributed to the understanding of population dynamics in the region: first using uniparental markers, particularly mitochondrial DNA in a much larger proportion; and more recently including genome-wide data for ancient individuals. In this work, we revise these studies considering three themes: (1) the first stages of migration into the region; (2) the diversification and interactions of populations during the Middle and Late Holocene; and (3) the link between present-day and ancient populations. While genetic evidence from the early peopling stages is either absent or scarce, making it difficult to evaluate the relative contributions of early South American lineages in the first Patagonian populations, evidence from later periods (from Middle Holocene onwards) is consistent with a single migration wave with founding events and genetic drift acting on small groups during their migration southward. After the initial occupation, the population dynamics seem to have been characterised by the relative isolation of different groups, leading to their differentiation. While there is evidence of some degree of gene flow between groups, the genetic structure in the region is generally associated with geography, subsistence systems, and languages. After European contact, paleogenetic data supports a relative genetic continuity in the region. We finish this review with a fourth theme in which we reflect on the current state and direction of the field in Patagonia, highlighting research lines that will benefit from the implementation of state-of-the-art paleogenomic approach, as well as legal and ethical considerations that would allow to move forward into a more collaborative and inclusive field.
Keywords
Paleogenetics, ancient DNA, uniparental lineages, genome-wide data, South America, Human Population Genetics, Demography
1. Introduction
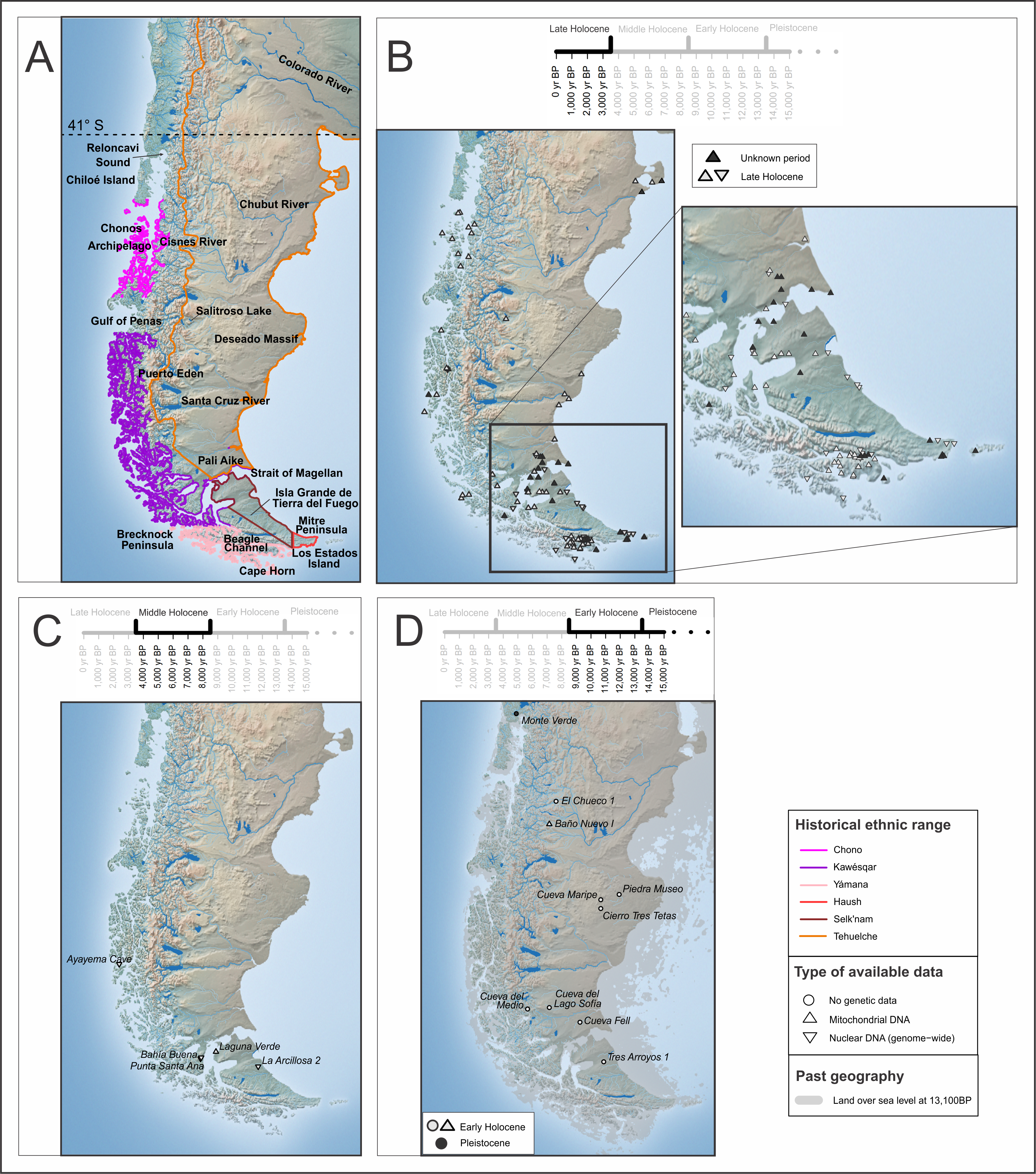
The population history of the Americas has been a matter of intense debates for decades with some regions occupying a more protagonist role. One such region has been Patagonia, where the evidence of early human occupation, its geographical and environmental diversity, and characteristics of human adaptations, has been a subject of continuous interest. Described as the “end of the world” and characterised by its harsh environment, Patagonia has been a centre of attraction for centuries. The region comprises the southern parts of Chile and Argentina (Figure 1A). However, its northern boundary varies between the western (Chile) and eastern (Argentina) portions following geo-political and environmental criteria. In geographic terms, the Andes are one of the most relevant features of the region, influencing the continental watershed (i.e., Pacific, and Atlantic) and in many cases indicating the international boundaries. The Pacific slope or western Patagonia extends from the 42ºS (Reloncaví Sound) to the south, reaching the Cape Horn area (~56ºS). Meanwhile, the north boundary of the Atlantic slope or eastern Patagonia is located south of the Colorado river (36ºS near the Andes and 39ºS in the coast), including the present-day provinces of Neuquén, Río Negro, Chubut, Santa Cruz, Tierra del Fuego and the southern part of Buenos Aires Province in Argentina [1]. In this review we focus on the region south of 41°S, considering the northern limit of Chilean Patagonia and extending it to the Atlantic coast (Figure 1). Therefore, we exclude northern Patagonia Argentina as its population dynamics are more closely related to central Chile and the Argentinean Pampas (Figure 1) [2–4].
Focusing on the characteristics of the human occupation of the region, several differences have been highlighted through archaeological, ethnographic, and ethno-historical sources, reflecting the cultural diversity in Patagonia. Within the archaeological record, there is evidence of changes in the use of the space and resources, interpreted as technological transitions [5]. Meanwhile, since the 16th century, populations in the region were characterised according to their subsistence strategies in maritime or terrestrial hunter gatherers, as well as some groups with mixed economies (see Box 1 for a brief contextualisation of the post-European contact period). Within maritime groups, which occupied most of western Patagonia, three major groups were identified: Chono, in the region south of Chiloé island to the Gulf of Penas; Kawésqar, immediately south of Chono until the Brecknock Peninsula; and Yámana, south of Brecknock Peninsula and on the Beagle channel shores (Figure 1A) [6–8]. In Tierra del Fuego, two other groups were recognized: Selk’nam, a terrestrial hunter gatherer group occupying most of the island; and Haush (or Manek-enk), characterised as a mosaic as they share features with terrestrial and maritime hunter gatherer groups. Both groups share the same language group [9], symbolic system [8], and the absence of navigation technology. Moreover, Haush is sometimes considered a sub-group within Selk’nam [8,10]. However, their high reliance on marine resources, evidenced by stable isotopes data [11], resemble maritime groups. Finally, Tehuelche populations had a terrestrial economy and occupied a vast territory in the continental Patagonia, north of the Magellan Strait. Some internal divisions among Tehuelche have been proposed from a linguistic perspective [9,12]. In particular, the groups inhabiting the south of Santa Cruz River name themselves Aónik-enk. It is worth mentioning that Tehuelche, Selk’nam, and Haush belong to the same Chon language family [13]; while Chono, Kawésqar and Yámana languages are described as isolated, albeit sharing a common origin [14]. All these groups maintained a nomadic hunter gatherer subsistence system by the time of contact with western society (see Box 1).
Box 1. Historical overview of European contact and national states territory incorporation. The available evidence from the historical period, considering this as the period from which written records of the region were done, suggest that the history of the Patagonian territories that are currently within the Argentinean and Chilean borders was similar in several ways. Europeans became acquainted with this region as part of the Magellan expedition in 1520. They started navigating through the area with the purpose of finding a path between the Atlantic and Pacific oceans, with sporadic encounters with local populations, leaving us topography names that reflect the kind of emotions that these lands inspired in them: e.g., “Puerto Hambre” (Port Hunger), “Bahia Desolación” (Desolation Bay), “Última Esperanza” (Last Hope). Scientific expeditions, aiming to describe the geographical, environmental, and demographic features of the area, did not start until the 18th century. Most of the early depictions of the indigenous populations made by European explorers described them as seemingly poorly adapted to the extreme cold and harsh conditions of the region [19]. Later on, during the second half of the 19th century, permanent settlements were established as part of religious missions with the goal of gathering indigenous people and imposing European culture, for example, Anglican in the Beagle Channel and Salesian in northern Tierra del Fuego and the western Archipelagos [20]. Subsequently, the Chilean and Argentinean nations undertook the appropriation of Indigenous territories, resulting in the confinement of local populations in religious missions. This phenomenon is particularly evident in the Argentinean Patagonia with the so-called “Conquest of the Desert” [21]. Similarly, on the Chilean side, the State’s interest on the industrial development of the region led to the establishment of several livestock (sheep) ranches or estancias (a phenomenon also observed in the Argentinean Patagonia), as well as to the exploitation of marine resources, with the corresponding immigration of Chilean colonists [22,23]. Along all this period, Indigenous communities were severely affected by diseases, wars, forced labour and relocations, witnessing a significant population decrease [24–26] since the establishment of the binational frontiers in 1902 as the result of the Pacific war (1881), both States have historically failed to recognise and validate the presence and ancestral rights of Indigenous Peoples [27]. In the 1990s, movements for recognition, rights and equality became publicly visible [28], and acquired greater strength in the 2000s, as a result of re-emergence processes of Indigenous Peoples wrongfully considered extinct [29–33]. Since 1994, the Argentinean Constitution recognizes the ethnic and cultural pre-existence of Indigenous Peoples, as well as the legal status of their communities, and the community possession and ownership of the lands they traditionally occupied (Art. 75 Inc. 17). Meanwhile, on the Chilean side only two Indigenous groups, Kawésqar and Yámana, were initially recognized by law (law Nº 19,253, 1993), and only recently (2023) Selk’nam was included. However, the creation of natural protected areas and exploitation of marine resources has been in conflict with the recognition of the traditional territories of the different communities in the region [22].
In this work, we provide a review of the population history of Patagonia, emphasising the archaeological and genetic evidence. In terms of genetic research, there have been multiple studies in the context of continental-wide initiatives as well as regional approaches, being a region intensely studied. To date, maternal lineage has been characterised and genome-wide data has been generated for ~300 individuals from the late to early Holocene (last 13,500 years before present; yr BP), respectively, most of which dated to the Late Holocene (last 3,500 yr BP; see Figure 1 for a timeline). While research across this sub-region has not been geographically or temporarily even (Figure 1), there is evidence suggesting shared cultural and biological features in the area, supporting the need of its consideration as a whole. We organise this review considering four major themes: (1) the first stages of migration into the region; (2) the diversification and interactions of populations during the Middle and Late Holocene; (3) the link between present-day and ancient populations; and (4) general considerations of paleogenetic studies in the region and future research directions.
2. First stages of human migration
Patagonia was initially occupied by small groups of hunter gatherers with different ranges of mobility [34]. This generated a dispersed archaeological record, making the paleodemographic study of these groups a challenging matter [35]. It has been suggested that the occupation of this region took place in a multidirectional process, dependent on environmental changes and the availability of resources [36]. This would have occurred in three stages: (i) exploration of the uninhabited territory, during which no evidence of human burials is expected to be found; (ii) colonisation, understood as the consolidation of groups in specific areas; and (iii) effective occupation of the space, in which every desirable territory is used [37].
The access point(s) of the first settlers to this region is particularly interesting given that it represents the end of the global dispersal path of Homo sapiens [36]. In western Patagonia, it has been suggested that the initial exploration stage would have taken place soon after the retreat of the glaciers. While there is no genetic or bioanthropological evidence associated with individuals from this early stage of human occupation, the Monte Verde site, dated to 14,500 cal yr BP (calibrated years before present) [38–41] in north-western Patagonia, would be a key evidence supporting this hypothesis [38,42], being the oldest record within the Southern Cone of South America (which encompasses present-day Argentina, Chile, Uruguay and southern Brazil). Archaeological evidence also suggests the presence of human populations in eastern Patagonia in the beginnings of the Pleistocene-Holocene transition (13,500 yr BP), probably in the early stages of occupation [43,44], considering the archaeological localities of Piedra Museo [45,46] and Cerro Tres Tetas [47] in the central Deseado Massif (Santa Cruz, Argentina) (Figure 1D). Further south, within the Magellan Basin, there are other early records of human activity in Cueva Lago Sofia [48], Cueva del Medio [49] and Cueva Fell [50], where fishtail projectile points and fires have been found dating up to 12,800 cal yr BP. On the present-day island of Tierra del Fuego, which was still connected to continental Patagonia [15], the oldest records of human activity come from fires located in the northern and central region of the island, from around 12,000 cal yr BP (Tres Arroyos 1 site) [51–53].
Around 10,000 yr BP, i.e., towards the end of the Early Holocene (13,500–8,500 yr BP), the colonisation stage would have begun, when the presence of populations is visible in the archaeological record, for example at the site El Chueco 1 (Cisnes River valley, central Chilean Patagonia) and Baño Nuevo-1 [40,41,54–56]. Notably, human burials have only been dated after this stage had begun, as to the oldest human remains reported are dated to ca. 10,000 cal yr BP (at Baño Nuevo-1 site; [41]). Several paleoenvironmental studies suggest that, at the beginning of the Late Holocene, the landscape conformation of the region was as it is today [57], and that the general improvement in climatic conditions could have favoured the expansion of the Patagonian populations that were already in a stage of effective occupation of the entire steppe [54,58,59].
Concerning the origins of the first inhabitants of the region, the first evidence comes from craniofacial morphometry studies which have proposed models with the arrival of two different populations to explain the variations observed in the ancient and present-day populations of the American continent [60–62]. Within this framework, Patagonian populations were described as “relicts” since their craniofacial morphology was associated with the earliest migration wave. Although initially supported by some mitochondrial DNA (mtDNA) studies [63], this hypothesis was challenged by other morphological investigations that attributed the observed phenotypic variability to the effect of random and non-random microevolutionary forces on a single migratory wave [64–67]. Later, mtDNA and, more recently, genome-wide studies, showed that the variation present in Patagonia is related to other South American lineages [68]. The absence of significant genetic differentiation between ancient individuals bearing these different morphologies, further support a common origin [69–71].
Ancient DNA studies of Y-chromosome have been carried out since the early stages of genetic research in the region [72–74], focusing on Short Tandem Repeats (STR) and the C to T transition at the DYS199 locus as a diagnostic polymorphism of Q-M3 haplogroup (also known as Q1b1a1a (ISOGG 2019)), a widespread lineage across the Americas. In all individuals from Patagonia yielding positive results, the lineage Q-M3 has been characterised. Given that the data from Y-STR is scarce, without enough overlapping markers, and low variability among individuals, the comparison between populations is difficult. Genome-wide studies did not change the picture substantially, since almost all the male individuals with enough coverage were found to carry the same Q-M3 lineage, or were otherwise classified inside the upper clade Q-M36 as the diagnostic position is missing [69,70,75,76]. The only exception is a Late Holocene individual from Mitre Peninsula with Q-Z780 [76], which is a low frequency clade spread across South America and Mexico. This lineage has a coalescent time slightly older than Q-M3 (19.3ky BP versus 15.4ky BP) and has been proposed as genetic evidence of an older peopling of South America [77]. The potential of whole Y-chromosome analysis for a fine scale population dynamic has yet to be exploited.
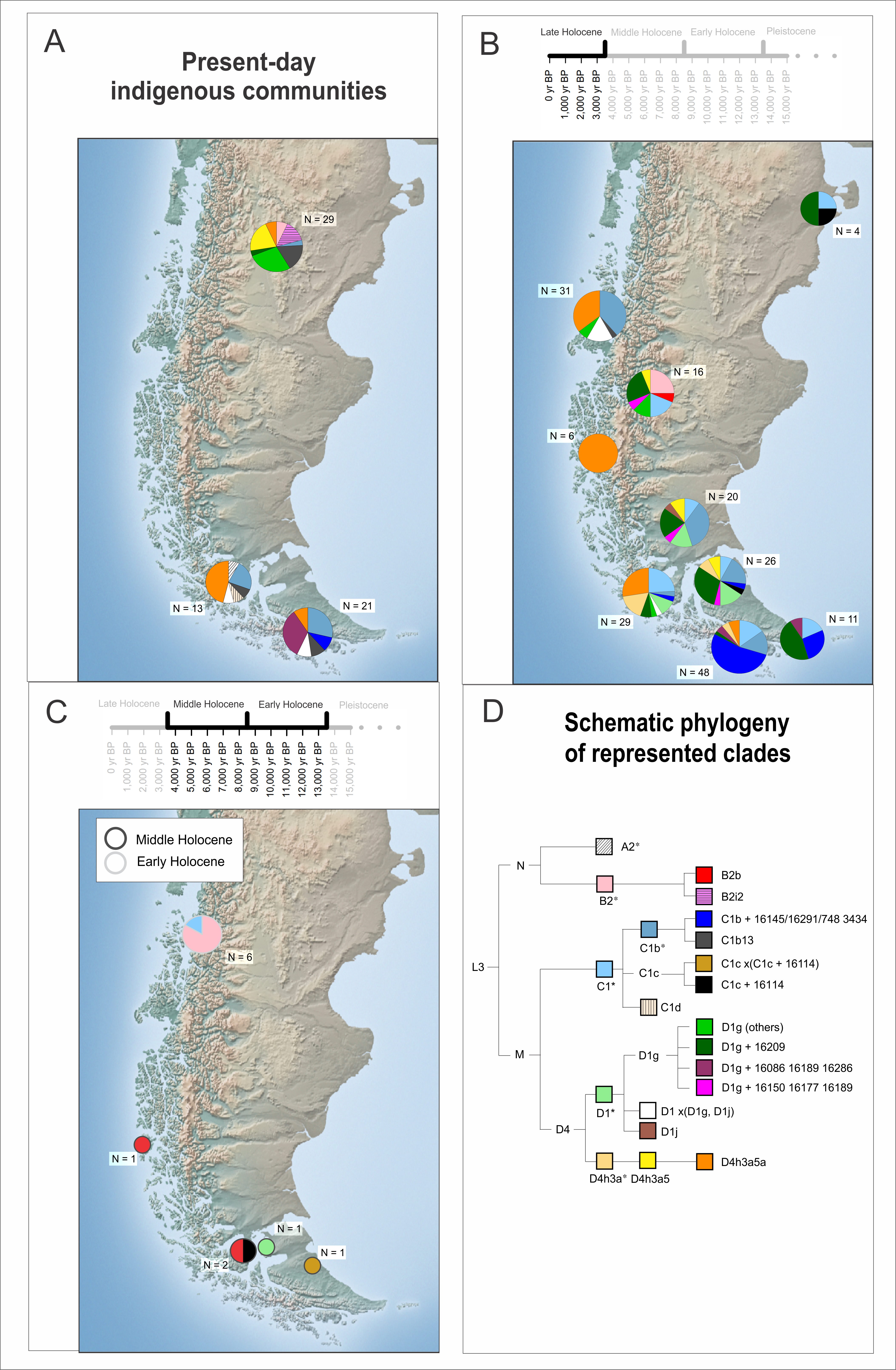
MtDNA studies have shown that Patagonia is the least diverse region in South America, with a prevalence of haplogroups C1, D1 and D4h3a5 towards the south, with B2 mainly restricted to the north at low frequencies [63,68,72,73,76,78–80] (Figure 2C and Table S1). This pattern is compatible with the effect of founding events and genetic drift acting on low population size groups during their southward dispersal. The earliest mitochondrial genetic evidence of Patagonia comes from the Baño Nuevo-1 site, dated to ca. 10,000 cal yr BP, and where 5 of the 6 individuals yielding enough DNA carry haplogroup B [41]. In addition, the lineage B2b has been characterised in two other locations from western Patagonian: the site Ayayema, dated to ca. 4,700 years BP [70] and site Bahía Buena dated to ca. 6,700 years BP [74], each with a single individual. Towards the Late Holocene haplogroup B2 is found at a frequency of 31% in Salitroso Lake basin [81,82] as well as in other areas of continental southern Patagonia (but with no peer-reviewed publication for those [83,84]). The early representation of haplogroup B suggests that this lineage was present in the first migratory waves towards the southern tip of the continent. Thus, its absence or limited presence in later populations of southern Patagonia (continental and insular) could be associated with an original low frequency and later loss by drift, sampling bias or gene flow associated with later migratory waves.
Moreover, mtDNA studies in present-day populations have been used to infer early entry routes and population movements. For instance, the distribution of the lineage D4h3a along the Pacific coast of South America has been suggested as evidence of the migratory route followed by the first populations [85]. In Patagonia, a regional sub-haplogroup (D4h3a5) has been described in ancient and present-day populations [78,85,86] (Figure 2 and Table S1). As D4h3a5 derives directly from the Beringia founding clade, this evidence suggests an early population arrival to the region probably following a Pacific route, in agreement with the archaeological evidence of early sites along the Pacific slope [87]. These early southward migrations may have been interrupted by ice sheets, which would have led to movements through the Atlantic slope [80]. Meanwhile, evidence from the Atlantic coast is still very scarce [88], which may be due to the great variation in its coastal geomorphology through time, especially during the Pleistocene-Holocene transition, when the deglaciation of the Cordilleran ice sheets occurred [36,89–93]. The geomorphology on the Atlantic and Pacific coasts (flat and steep, respectively) implied a different reconfiguration of the coastlines on both sides (Figure 1D), making it more likely that the earliest evidence of human occupation is currently underwater for the eastern settlement route (in the submarine plains) than for the western one (in submarine abrupt waterscape).
Considering the evidence from genome-wide data, ancient and present-day individuals from Patagonia are part of the major South Native American (SNA) lineage, first characterised in Anzick-1 individual from Montana (USA) and linked to the Clovis archaeological culture [69–71,75,76,94]. Several SNA lineages have been described so far, and the links among them and their relative contributions into the different regions of South America remain to be disentangled [70,71,95,96]. The evidence of a putative replacement across South America of a first lineage (SNA1) associated with Anzick-1 by later migration waves [71] is challenged as new genomes are generated and new demographic models are suggested [96,97]. Later migration waves (i.e., not related to SNA1), have been assigned to SNA2 lineage, associated with individuals linked to the Western Stemmed Tradition [98]. However, the term SNA2 is used to describe lineages that have different origins (e.g., see the models reported in [95,96]). These pulses of population influx appear to have a heterogeneous genetic legacy across the subcontinent [70,71,95], and their importance in the demographic history of Patagonia remains unclear. However, a recent study proposed that the SNA2 lineage, associated with ancient individuals from North America (Spirit Cave site), would have contributed directly to all Patagonia populations, in contrast to an early wave associated to Anzick-1 (SNA1) [95]. The presence of other early lineages (i.e., population Y or Australasian-related ancestry) or later migrations (i.e., migrations associated to Mixe and Channel-Island related ancestries during the Middle Holocene) have not been found in Patagonia [70,71].
3. Diversification and interactions among human populations during the Middle and Late Holocene
Through the analysis of uniparental markers (mtDNA and Y-chromosome) and genome-wide data, we have also learned about the genetic diversity and structure of the populations that inhabited Patagonia during the Middle and Late Holocene. As mentioned before, in general terms human groups from the region are characterised by a reduced genetic diversity compared to other populations in the Americas. This was initially suggested by early mtDNA research, highlighting the lack of certain mitochondrial lineages, particularly A and B haplogroups [63]. Although later work has found evidence of B2 mtDNA lineages in the region, their frequency as well as their geographical and temporal distributions are quite restricted (Figure 2). In addition, the identification of these lineages has been possible only after a significant increase in the sample size and geographical coverage. The overall low uniparental genetic diversity [63,72–74,76,78,99], together with the reported excess of short Runs of Homozygosity [75] and the low conditional heterozygosity [76] from genome-wide data, support a reduced effective population size. This scenario is compatible with the founder effect(s) during the peopling of the region, followed by isolation and persistent small group sizes. However, the data produced so far has not allowed to estimate the demographic changes across time. Particularly, the question of effective population size during the initial peopling and later periods remains open.
Despite the overall low uniparental genetic diversity, since 2015 genetic studies have been providing insights into population dynamics and differences within Patagonia, especially during the Middle and Late Holocene. A broad comparison, performed by Crespo et al. [100], compiled the data available in the literature for the mtDNA Hypervariable Region I (HVRI), finding significant differences between three main regions: north, south, and insular Patagonia. Although this analysis groups together individuals usually considered separately based on their cultural features (e.g., north Tierra del Fuego and Beagle Channel), it provides a more comprehensive approach and has the strength of supporting the heterogeneity of populations in Patagonia from a molecular perspective. Differences in mtDNA diversity has also been evaluated in a later work by comparing individuals according to their inferred subsistence strategy: maritime, terrestrial and mixed economies, finding that maritime groups are more diverse than the terrestrial ones, while no significant differences has been found between terrestrial and mixed economy groups [74].
Further differences have been evaluated comparing the mtDNA variation between individuals from the Patagonian Archipelagos. De la Fuente et al., [73] compared the mtDNA diversity using the HVRI and II of ancient individuals dating to the last 2,000 years. While previous research suggested the presence of only C and D lineages, this work showed significant differences between maritime hunter gatherers from two regions (Figure 2B): south of the Beagle Channel (with C1 lineages at high frequency) and the western Archipelagos between Brecknock peninsula and Puerto Edén (with D4h3a lineages at high frequency). More recently, the mitochondrial DNA variation during the Late Holocene of the northern part of the western archipelago, in particular the Chono archipelago, was analysed. Moraga et al., [78] found significant differences between ancient individuals from the Chono archipelago and the ones described further south, supporting the presence of a distinctive group in this area. However, the authors also described some shared lineages within maritime hunter gatherers from other areas of the Patagonian archipelago. Moreover, the analysis of mitogenomes allowed them to describe a monophyletic derived clade, D4h3a5a1, which has been almost exclusively found in maritime hunter gatherers across the western Archipelago. Since this sub-lineage has not been identified in terrestrial groups [75,76], it supports a deep shared history for most maritime groups or some level of interaction among them [78], which is further supported by genome-wide analysis [75,76]. However, the ancestral D4h3a5 is observed in present-day Tehuelche [80], and in individuals from the Late Holocene in south-eastern Patagonia [81,86] and the Late Holocene (including historical) from north Tierra del Fuego [76,101]. This difference between terrestrial and maritime haplotypes, with more derived ones in the latter, was interpreted as evidence supporting a local origin of maritime populations [73,78,86].
The genetic data of ancient individuals from mainland Patagonia is more limited than in the archipelagos, since the available information comes mainly from mtDNA HVR of a handful of Late Holocene individuals from south Santa Cruz province in Argentina (south-eastern continental Patagonia) [86,102,103], four individuals from the coast of Chubut (north-eastern continental Patagonia) [72,104], and 16 individuals from Salitroso Lake basin (north-western Santa Cruz) [81], where two mitogenome are available [82]. The lineages found are those typically characterised in Patagonia (C1, D1g and D4h3a5), with the exception of one individual carrying the haplogroup D1j, which was interpreted as an inland or Atlantic coast migration reaching south Patagonia [102], as well as B2 haplogroup present in a third of Late Holocene individuals from Salitroso Lake basin [81] (Figure 2). Moreover, based on shared derived haplotypes, some level of genetic affinity between Salitroso Lake, south-eastern continental Patagonia and north Tierra del Fuego is detected [81], as well as between south-eastern continental Patagonia and Tierra del Fuego island [103]. Altogether, genetic diversity indicates that Tierra del Fuego populations may not have been completely isolated from mainland Patagonia, albeit the island has been separated from the rest of continental Patagonia since around 8,000 yr BP when the rise of sea level established the Strait of Magellan as a geographical barrier for humans [15,44,105,106].
Genome-wide approaches have been implemented in Patagonia as part of broad continental contexts [69,70] and regional questions [75,76]. To date, 39 ancient genomes have been recovered from different temporal, geographical and archaeological contexts, although most individuals (n = 35) date to the Late Holocene period (Figure 1). Similar to the analysis using uniparental inherited markers, genome-wide analyses aim to describe the unique genetic variation of Patagonian populations and the dynamics within the region that had led to the differences between groups. A first sign of differentiation contributing to the population structure is associated with the development of a maritime tradition in western Patagonia about 7,000 years BP. The earliest human individual clearly associated with this tradition is from the archaeological site Punta Santa Ana and dated to ca. 6,600 BP. The genome of this individual shared the highest genetic drift with other Middle and Late Holocene individuals from Patagonia, supporting the long-term continuity in the region [70,76]. Another Middle Holocene individual, from north Tierra del Fuego dated to ca. 5,800 BP (La Arcillosa 2), had primarily a terrestrial diet and showed a genome-wide signal similar to Punta Santa Ana individual, suggesting a local origin of maritime adaptations from terrestrial hunter gatherers [76]. In fact, when comparing their genetic affinities against other ancient and present-day populations from Patagonia, they both are equally related to later populations [76]. However, from 4,700 years BP there is evidence of genetic differentiation separating individuals with a maritime and terrestrial subsistence (or economy). This date is given by the individual from the site Ayayema, in central western Patagonia, the second oldest individual associated with a maritime tradition and whose genome has been recovered. This first differentiation does not seem associated with external migrations from outside Patagonia, but rather with population movements along the western Archipelago. In addition, a second migration into south Patagonia has been inferred from whole genomic data which did not involve population replacement but contributed substantially to all Late Holocene individuals. The source of this population is primarily ascribed to central Chile, but this is probably due to the lack of whole genome data for the intermediate regions [76].
Another topic of analysis has been the association between genetic and fine-scale geographic and cultural differences. Within Late Holocene groups (<3,500 BP) there is some degree of genetic structure among groups following the traditional territories attributed to the populations: Kawésqar and Yámana in western Patagonia, and southern Tehuelche (Aónik-enk), Selk’nam, and Haush to the east [73,75,76]. While there is also evidence of a gradient of gene flow between neighbouring populations following the coastline around 2,200–1,200 years BP [76], this gene flow was not enough to blur genetic differences among groups associated with different subsistence strategies and languages. A particular case is represented by the inhabitants of Mitre Peninsula, who from a genetic perspective are a mixture between north Tierra del Fuego and Beagle Chanel populations [76], consistent with what was observed with mtDNA [107].
Overall, genetic evidence has supported the differentiation and structuring of Patagonian populations associated with geographical and cultural (subsistence strategy and language) features. The landscape of Patagonia, especially on the western side, certainly contributes to the relative isolation of the different groups through time, but with some degree of gene flow mostly between neighbouring groups in Tierra del Fuego.
4. Exploring the relationship between early and present-day populations
The genetic data obtained from both modern (Box 2) and ancient individuals provides the opportunity to better understand the effect of colonisation on the genetic variability in the region. Considering uniparental inherited markers, to date there are no specific studies about the composition of paternal lineages in Indigenous Peoples of south Patagonia, but 5 to 12% of the studied individuals form the general population in the region carry an American paternal lineage [108–112], which is consistent with what is observed in the Southern Cone of South America [111,113,114]. In contrast, almost all studied present-day individuals from Indigenous communities in the region carry Indigenous maternal lineages (A2, B2, C1b, C1d, D1 and D4h3a; Figure 2A) [80,99], also in agreement with the results reported for other Southern Cone of South America Indigenous or rural populations [115–120].
Box 2. Indigenous recognition and its impact in genetic research through time. The changes concerning the recognition of indigenous communities (Box 1) have had a counterpart in paleogenetic analyses. The first ancient DNA studies of human populations from Patagonia described them as “Extinct Aboriginal Tribes” [63,72,121], reflecting the invisibilization of Indigenous Peoples at the time. However, later, and most probably as a direct consequence of the movements for the recognition of Indigenous Peoples, this preconceived idea of extinction has been abandoned. Furthermore, genetic data from present-day communities in south (Yámana), west (Kawésqar) and north (Tehuelche) Patagonia have been generated in the 2010s aiming to evaluate the diversity of these populations through uniparental markers [80,99], and later adding nuclear genome-wide markers in western Patagonia [75,122]. While the specific research aims and perspectives about cultural and biological continuity through time varies in the scientific literature [63,72,75,76,107,121], most authors seem to implicitly or explicitly agree regarding the continuity of these populations, at least from the Middle Holocene until European colonisation. Moreover, during three decades, researchers have been attributing ethnonyms to pre-contact individuals according to the spatial distribution of the different Peoples encountered by Europeans when they occupied the region (see [123] for a discussion on the use of ethnonyms in paleogenetics).
More in detail, it is interesting to highlight the presence of shared mtDNA clades or haplotypes between present-day [80] and ancient individuals [73,74,107,124] from the same geographic area, such as the lineage D4h3a5a reaching 46% in present-day Kawésqar, and C1b+16291-195 and D1g+16086-16189-16286 with a combined 43% in Yámana. However, there is evidence of a few clades (e.g., C1b13 and A2) that may be the results of historical migrations into the region [79,107]. Regarding present-day Tehuelche, the data from ancient individuals from mainland Patagonia is still scarce and/or does not have enough resolution for a more detailed comparison. Notwithstanding, the two sub-haplogroups described in ancient individuals from this specific geographic area (D1g + 16209 and D4h3a5 without the reversion in 16301) has been registered in present-day individuals (Figure 2) [81,86].
Considering autosomal variation, members of Indigenous communities in the region exhibit high proportions of Indigenous ancestries, albeit usually lower than in other South American Indigenous populations [75,122]. When focussing only on the Indigenous genetic ancestry, a strong genetic continuity is observed between present-day and ancient individuals. Furthermore, ancient individuals from southern and southwestern archipelagos are closer to present-day Yámana and Kawésqar than to any other present-day population [75]. These findings have been later reinforced when analysing individuals covering a larger geographic range [76]. De la Fuente et al., [75] also showed that there was a high geographical correlation, where ancient individuals were genetically closer to either present-day Yámana or Kawésqar depending on their proximity to the territories traditionally associated with those populations. This study also showed that modern populations from Chiloé and central-south Chile are genetically closer to modern Kawésqar than to modern Yámana. This cannot be attributed to historical population movements, as those populations (Chiloé and central-south Chile) are also closer to ancient individuals from the southwestern Archipelago (the geographic region of Kawésqar) than to other ancient Patagonian populations [76]. Moreover, this cline of genetic affinities may have originated from a decreasing intensity of gene-flow from central Chile into southern regions as mentioned above [76]. Disentangling the demographic processes that originated this cline may be possible as more ancient individuals from different Patagonian regions are analysed, as well as individuals from the potential source locations of this gene flow.
Finally, Luisi et al., [125] showed that modern individuals from general populations in southern Argentina exhibit the highest proportions of a genetic ancestry component mainly observed in ancient and modern Indigenous individuals from central Chile and Patagonia. This result suggests that, despite population movements occurring since colonisation, a specific genetic ancestry, which emerged several millennia ago through population divergence with other South American populations [76,95,125], remains nowadays highly represented in the region.
5. General considerations and future directions
As many other fields in science, paleogenetic research in Patagonia has not been exempt from subjective perspectives. For example, interpretations of the low genetic diversity in the region varies from early research considering populations from the region as “relicts”, isolated and confined to the southern end of South America as a consequence of successive migrations that pushed them southwards [63,72], to later interpretations taking the generally low diversity of Patagonian populations as the effect of genetic drift [68]; and finally more recent studies highlighting the interactions between groups and movements [75,76]. The isolation paradigm was more in syntony with the early views regarding the "extinction" of indigenous Patagonian populations, whereas the consideration of more complex scenarios contributes to highlighting the role of the economical and socio-political factors (Box 1) at stake in their population decline [126].
In addition, researchers working in this field should be aware of the social implications of the construction of the past, and pursue a more active engagement with the local communities starting from the design of the research projects [127], attending to local questions, needs and concerns. We encourage future paleogenetic projects in Patagonia to pursue a more collaborative approach with Indigenous organisations, today a very limited practice in South America [123], to address the erasure of Indigenous voices and the lack of co-construction of Indigenous histories. We recognize this is not an easy task and it should not be underestimated or romanticised as each Indigenous community has its own dynamics and internal discussions. Furthermore, the current scientific structure (e.g., funding agencies, journals) should favour and promote initiatives that include Indigenous participation. Finally, we would like to stress the importance of considering the legal framework usually involved in paleogenetic research, including heritage management regulations and Indigenous rights [128].
While writing this review, we were increasingly aware of the heterogeneity of the analytical units across the different studies. Different grouping criteria have been used, such as the geographic ranges of 19th century ethnic groups (i.e., ancient Kawésqar, ancient Yámana and ancient Selk’nam), the subsystem system (i.e., maritime vs. terrestrial groups), or geography (i.e., north, central, and south Patagonia). Indeed, some cautions have been raised against the use of historic ethnonyms that could be masking internal divisions and/or diachronic changes [107]. Actually, the generation of artificial sets of individuals to be considered as evolutionary units is a common issue in ancient genetic studies. Those evolutionary units are flexible and dynamic and are somewhat arbitrarily built according to the analytical framework of the study. Special attention is therefore needed when comparing the results across different studies and to the dynamic meaning of the names used [129].
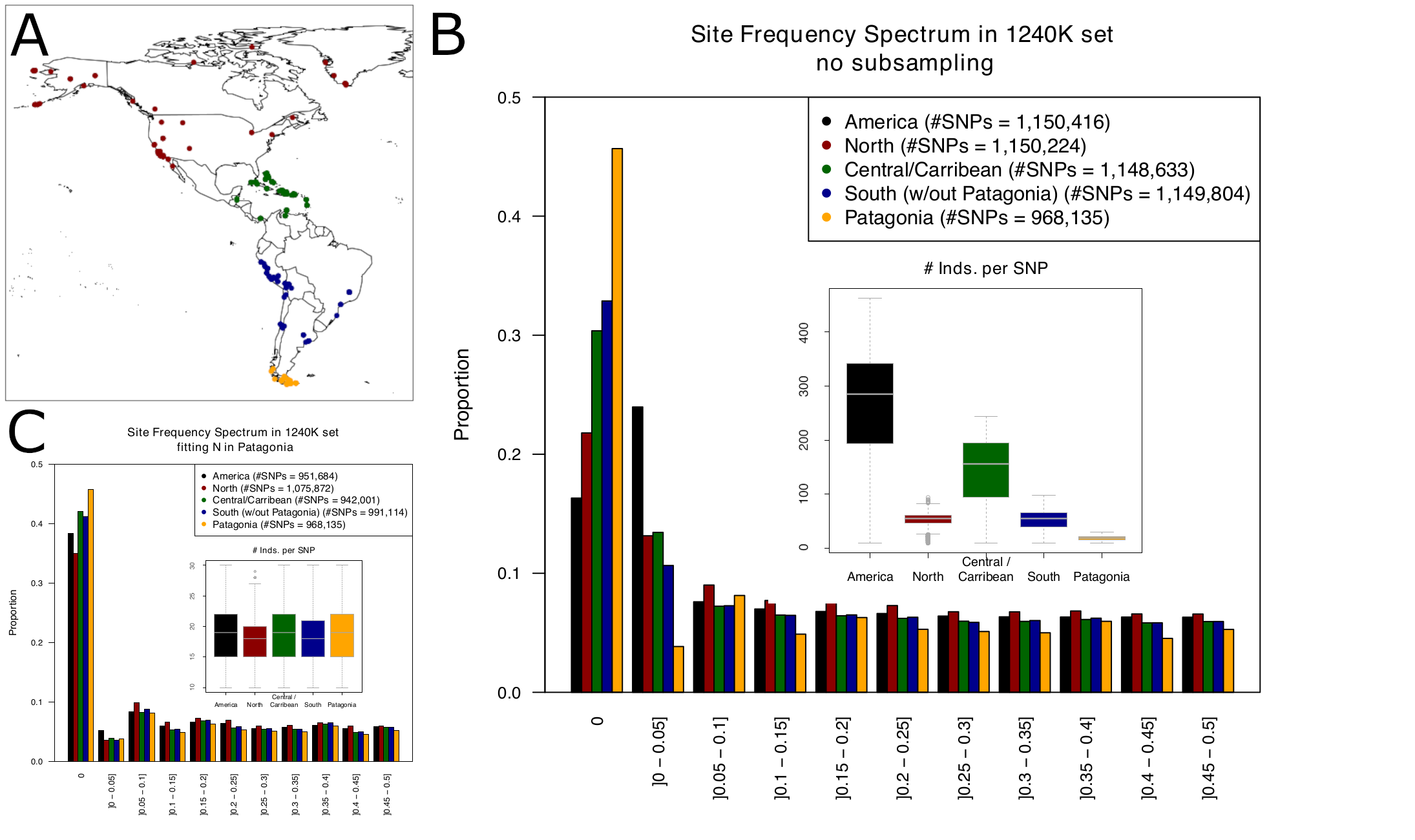
From a more technical perspective, when working with ancient DNA, special attention should be put on reference biases introduced by mapping post mortem damaged DNA to the reference genome, as these biases vary by coverage, read length and sequencing depth, thus distorting allele frequency estimates [130]. Most of the human paleogenomic data generated in Patagonia has been limited to a SNP panel of ~1,240k positions (hereafter 1240K SNP array) [131–133]. This approach has been highly favoured by many members of the paleogenomic community [134], mainly due to the alleged economical advantage and the increased DNA recovery rate in samples with poorly preserved DNA. However, while it is not clear this benefits research groups equally [135], there has also been increasing concerns about possible biases in the data compared to shotgun sequencing, including a strong reference bias [136]. Additionally, a systematic assay bias that induces higher genetic similarity between individuals enriched with the same kit has been suggested [137]. These biases may lead to erroneous conclusions when the target populations are highly genetically differentiated from the populations used in the ascertainment scheme to build the capture array, which is the case with Patagonian populations. In particular, ~46% of the sites from the 1240K SNP array are monomorphic in Patagonia, most probably because of the combined outcome of ascertainment bias and sequential founder effects that led to allele fixation in this remote part of the world (Figure 3). In other words, almost half of the data generated with the 1240K SNP array is not informative to analyse local demographic processes in Patagonia. This figure is better in other regions of the Americas where more individuals have been sequenced to date (Figure 3B), but the differences remain very narrow when reducing the sample size (Figure 3C). Now that more data is being generated for this region, it may prove useful to design new arrays with specific ascertainment schemes that encompass the genetic diversity of the region. Alternatively, a shotgun sequencing approach may be more suitable for capturing the genome-wide diversity in Patagonia and perform a wide range of analyses not feasible otherwise, such as estimating divergence times between human groups, demographic dynamics over time, and accurate estimations of kinship among individuals, all being central questions in archaeology. For instance, high coverage shotgun sequencing for ancient individuals before and after contact with European society would allow estimating the intensity of the expected decrease in genetic diversity, and thus inferring the reduction in population sizes that occurred at that time. This analysis has already been performed in North America through exome sequencing and limiting the analyses to a reduced number of synonymous (putatively neutral) variants [138]. Thus, shotgun sequencing data from Early and Middle Holocene individuals may provide useful insights into the population size changes during the three stages of occupation of the region (exploration, colonisation, and effective occupation as proposed by Borrero et al., [37]).
Furthermore, inferring genetic kinship relationships would provide a useful source of information to better understand micro scale mobility patterns among different nearby geographic areas, or long distance migrations [139], as well as the social structure within each group [140]. However, reaching robust kinship estimations implies to overcome several issues such as the relative reduced effective population size in the region, the relative small sample sizes that complicate the calibration of the underlying models - required for instance in READ [141] or Kin [142], or the lack of modern reference population to estimate representative allele frequencies - required for instance for lcMLKin [143] and TKGWV2 [144]. Generating shotgun sequencing genomes at >1X may facilitate kinship estimations leveraging robust methods such as IBSrelate [145]. In addition, with shotgun sequencing, paleogenomics studies could also retrieve DNA from microorganisms (including pathogens), elements from the diet or the environment, thus opening the opportunity to gain insights on the health, diets and plant usage for medicinal purposes in past populations [146]. By analysing the complex interplay among lifestyles, past human migrations, and diseases in different regions, we could improve our understanding of the dynamic nature of past human populations, hopefully breaking with the still wide-spread vision of them being “frozen in time” [147].
Although it has not been studied in depth, height of Patagonian individuals has also been pointed out as an interesting research theme, as it shows pronounced differences between maritime and terrestrial hunter gatherers of the region [148]. Moreover, this trait is a good candidate for phenotypic adaptation in the region since it is a complex trait with above-average heritability [149,150], has evidence of polygenic adaptation in Europe and Africa ([151]; but see [152]), and it is expected to be higher in populations inhabiting colder environments (or with fewer cultural adaptations to cold) for thermoregulation according to the Bergmann’s rule [153]. Genetic evidence would potentially provide useful insights to disentangle the different forces at stake in other morphological differences observed among groups [154–156]. For instance, comparing morphologic and genetic differentiations among worldwide populations, Zaidi et al. [157] concluded that nose shape may have been driven by local adaptation to climate to some extent. Actually, recently, a potentially adaptive Neanderthal introgression occurring almost exclusively on an Indigenous chromosomal background seems to increase nasal height [158].
6. Conclusions
Studying genetic differences between nearby areas in Patagonia has been possible even with low-resolution studies such as mtDNA haplotypes, thus contributing to our understanding of the populations’ history in the region. Incorporating genome-wide data, and the increasing representation in time and space of individuals have proven to be essential to address regional archaeological questions and to get a better understanding of the deep history of human populations. However, many questions remain open, and to which paleogenomics could provide useful information.
First, we still lack a robust demographic model that encompasses the evolutionary links with other regions of South America through time to explain (i) the early peopling of the region, (ii) how north-to-south genetic clines emerged during the Late Holocene, and (iii) how population sizes changed across time, among others. A better representation of continental Patagonia and high-resolution genomic data through deep shotgun sequencing may be determinant to calibrate many of these features. Beyond demography, paleogenomics may also be useful to tackle open questions on the social structure among and within groups, their lifestyle, mobility patterns, health, and biological adaptation to their environment. Finally, integrating evidence from paleogenomics with that derived from other disciplines, including the perspectives from Indigenous Peoples, will not only enrich the knowledge produced by our research, but will also facilitate a more socially responsible science.
Supplementary Materials
The following supplementary materials are available at: HPGG2404010003SupplementaryMaterials.zip
Declarations
Ethics Statement
Not applicable.
Consent for Publication
Not applicable.
Availability of Data and Material
Not applicable.
Funding
JM was funded through Consejo Nacional de Investigaciones Científicas y Técnicas, Grant/Award Number: PIP 2021-2023 N° 11220200103037CO. VA was funded through Consejo Nacional de Investigaciones Científicas y Técnicas and Fundación Científica Felipe Fiorellino. PL was funded through Consejo Nacional de Investigaciones Científicas y Técnicas.
Competing Interests
The authors have declared that no competing interests exist.
Author Contributions
All authors contributed equally to the design, writing, and editing of this manuscript.
Acknowledgement
We are grateful for the comments provided by the anonymous reviewers and editors, and their dedication to improve the quality of this manuscript.
References
| 1. |
Coronato AMJ, Coronato F, Mazzoni E, Vázquez M. The Physical Geography of Patagonia and Tierra del Fuego. In: Rabassa J, editor. Developments in Quaternary Sciences. Volume 11. Amsterdam: Elsevier; 2008. p.13-55.
[Google Scholar]
[CrossRef]
|
| 2. |
Barrientos G, Pérez SI. La expansión y dispersión de poblaciones del norte de Patagonia durante el holoceno tardío. Contra viento y marea/Civalero [Internet]. 2004 [cited 2024 Jan 22]; Available from: http://naturalis.fcnym.unlp.edu.ar/id/003863.
|
| 3. |
Nacuzzi LR. Los grupos nómades de la Patagonia y el Chaco en el siglo XVIII: identidades, espacios, movimientos y recursos económicos ante la situación de contacto. Una reflexión comparativa. Chungará (Arica). 2007;39:221-234.
[Google Scholar]
[CrossRef]
|
| 4. |
Flegenheimer N, Miotti L, Mazzia N. Rethinking early objects and landscapes in the Southern Cone: Fishtail-point concentrations in the Pampas and Northern Patagonia. In: Graf KE, Ketron CV, Waters MR, Graf KE, Ketron CV, Waters MR, editors. Paleoamerican Odyssey. Paleoamerican Odyssey Conference Companion Book. College Station: Center for the Study of the First Americans, Texas A&M University; 2013. p.359-376.
[Google Scholar]
|
| 5. |
San Román M, Reyes O, Morello F, Torres J. Archaeology of maritime Hunter-Gatherers from Southernmost Patagonia, South America: discussing timing, changes and cultural traditions during the Holocene. In: Bjerck HB, Breivik HM, Fretheim SE, Piana EL, Skar B, Tivoli AM, Zangrando AFJ, editor. Marine ventures Archaeological perspectives on human-sea relations. Scheffield: Equinox; 2016.
[Google Scholar]
|
| 6. |
Bridges EL. Uttermost part of the earth. London, UK: Hodder & Stoughton; 1951.
|
| 7. |
Emperaire J. Los Nómades del Mar (Traducción de Luis Oyarzún). Santiago: Ediciones de la Universidad de Chile; 1963.
|
| 8. |
Gusinde M. Los indios de Tierra del Fuego: Los Selk’nam (2 v.). Buenos Aires: Centro Argentino de Etnología Americana, Consejo Nacional de Investigaciones Científicas y Técnicas; 1982.
|
| 9. |
Viegas Barros JP. La familia lingüística tehuelche. Revista Patagónica. 1992;54:39-46.
[Google Scholar]
|
| 10. |
Saletta MJ, Fiore D. As time goes by. Cantidad de información, visibilidad e intensidad de registro de la tecnología shelk’nam-haush en el registro histórico-etnográfico producido por viajeros, misioneros e investigadores en Tierra del Fuego desde el siglo XVI al XX. [Internet]. 2017 [cited 2023 Nov 14]; Available from: https://ri.conicet.gov.ar/handle/11336/75341.
|
| 11. |
Yesner DR, Torres MJF, Guichon RA, Borrero LA. Stable isotope analysis of human bone and ethnohistoric subsistence patterns in Tierra del Fuego. J Anthropol Archaeol. 2003;22(3):279-91.
[Google Scholar]
[CrossRef]
|
| 12. |
Casamiquela RM. Rectificaciones y ratificaciones hacia una interpretación definitiva del panorama etnológico de la Patagonia y área septentrional adyacente. Bahía Blanca: Universidad Nacional del Sur; 1965.
|
| 13. |
Censabella M. Las lenguas indígenas de la Argentina : una mirada actual. Buenos Aires: EUDEBA; 1999.
|
| 14. |
Barceló JA, del Castillo MF, Mameli L, Moreno E, Videla B. Where Does the South Begin? Social Variability at the Southern Top of the World. Arctic Anthropol. 2009;46(1-2):50-71.
[Google Scholar]
[CrossRef]
|
| 15. |
McCulloch R, Morello F. Evidencia glacial y paleoecológica de ambientes tardiglaciales y del Holoceno temprano. Implicaciones para el poblamiento temprano de Tierra del Fuego. Arqueología de Patagonia: una mirada desde el último confín. 2009;119-133.
[Google Scholar]
|
| 16. |
Wickham H, Chang W, Wickham MH. Package “ggplot2.” Create elegant data visualisations [Internet]. 2016 [cited 2023 Nov 14]; Available from: https://citeseerx.ist.psu.edu/document?repid=rep1&type=pdf&doi=af53fd2f5b9e81b6edec0c13e1b3babd34bda399.
|
| 17. |
Lehner B, Grill G. Global river hydrography and network routing: baseline data and new approaches to study the world’s large river systems. Hydrol Process. 2013;27(15):2171-2186.
[Google Scholar]
[CrossRef]
|
| 18. |
Messager ML, Lehner B, Grill G, Nedeva I, Schmitt O. Estimating the volume and age of water stored in global lakes using a geo-statistical approach. Nat Commun. 2016;7:13603.
[Google Scholar]
[CrossRef]
|
| 19. |
Darwin C. On the origin of species: A facsimile of the first edition. with an introduction by Ernst Mayr. With a portrait and a bibliography. London: John Murray; 1860. 502 p.
|
| 20. |
Casali R. Contacto interétnico en el norte de Tierra de Fuego: primera aproximación a las estrategias de resistencia Selk’nam. Magallania. 2008;36(2):45-61.
[Google Scholar]
[CrossRef]
|
| 21. |
Rivas Maldonado J, Mora T, Rodriguez C, Emilson CR, Loyola Gómez Christian Y. Patagonia, territorio de los otros: consideraciones geográfico-políticas en la construcción de la nación Argentina [Internet]. 2015 [cited 2023 Nov 14]. Available from: https://www.redalyc.org/pdf/3477/347743079007.pdf.
|
| 22. |
Aylwin J, Arce L, Guerra F, Núñez D, Álvarez R, Mansilla P, Alday D., Caro L., Chiguay C., Huenucoy C. Conservacion y pueblos indigenas en la Patagonia Chilena. In: Castilla JC, Armesto JJ, Martínez-Harms MJ, editor. Conservación en la Patagonia chilena: evaluación del conocimiento, oportunidades y desafíos. Santiago: Ediciones Universidad Católica; 2021. p.495-531.
[Google Scholar]
|
| 23. |
Mandujano-Bustamante F. O Estado chileno ea Patagônia: conflitos territoriais. Bitácora Urbano [Internet]; 2016 [cited 2023 Nov 14]; Available from: http://www.scielo.org.co/scielo.php?pid=S0124-79132016000100010&script=sci_abstract&tlng=pt.
|
| 24. |
Casali R. Relaciones interétnicas en Tierra del Fuego: el rol de la misión salesiana La Candelaria (1895-1912) en la resistencia selk’nam. Universidad Nacional de Mar del Plata. Grupo de Estudios Sociales Marítimos. Revista de Estudios Marítimos y Sociales. 2013;5/6:105-117.
[Google Scholar]
|
| 25. |
Mandrini R. La Argentina aborigen: de los primeros pobladores a 1910. Buenos Aires: Siglo XXI Editores Argentina; 2008.
|
| 26. |
Martínez Sarasola C. Nuestros paisanos los indios [Internet]. 2011 [cited 2023 Nov 15]; Available from: https://www.repositoriointerculturalidad.ec/jspui/handle/123456789/32161.
|
| 27. |
Lenton DI, Delrio WM, Pérez PMV, Papazian AER, Nagy MA, Musante M. Argentina’s constituent genocide: challenging the hegemonic national narrative and laying the foundation for reparations to indigenous peoples. Armenian Review. 2012;53(14):63-84.
[Google Scholar]
|
| 29. |
Bompadre JM. (Des) Memorias de La Docta. De barbudos miscegenados a comechingones comunalizados: procesos contemporáneos de emergencia étnica en Córdoba. Córdoba: Universidad Nacional de Córdoba; 2016.
|
| 30. |
Bustos MA. La dimensión política de la identidad y los conflictos sociales. Confluencia. 2003;1(1):185-194.
[Google Scholar]
|
| 31. |
Di Biase ARS. Nosotros somos los verdaderos originarios”: la construcción de la identidad indígena en la pampa. Buenos Aires: Universidad de Buenos Aires; 2019.
|
| 32. |
Palladino L. Territorio, comunidad e identidad. El proceso de comunalización de los comechingones del Pueblo de La Toma, Ciudad de Córdoba (2008-2009) [Internet]. 2013 [cited 2023 Nov 14]. Available from: https://revistas.unc.edu.ar/index.php/cardi/article/view/7063/8136.
|
| 33. |
Ramos AM, Bompadre JM. Procesos de Memoria: Introducción [Internet]. 2018 [cited 2023 Nov 14]. Available from: https://ri.conicet.gov.ar/handle/11336/103720.
|
| 34. |
Borrero LA. Early Occupations in the Southern Cone. In: Silverman H, Isbell WH, editor. The Handbook of South American Archaeology. New York, NY: Springer; 2008. p.59-77.
[Google Scholar]
[CrossRef]
|
| 35. |
García Guraieb S, Goñi R, Tessone A. Paleodemography of Late Holocene hunter-gatherers from Patagonia (Santa Cruz, Argentina): An approach using multiple archaeological and bioarchaeological indicators. Quat Int. 2015;356:147-158.
[Google Scholar]
[CrossRef]
|
| 36. |
Borrero LA. El Poblamiento de La Patagonia: Toldos, Milodones y Volcanes. Buenos Aires: Emece Editores; 2001.
|
| 37. |
Borrero LA. Evolución cultural divergente en la Patagonia austral. Anales del Instituto de la Patagonia [Internet]; 1989 [cited 2023 Nov 14]; Available from: http://www.bibliotecadigital.umag.cl/handle/20.500.11893/957.
|
| 38. |
Dillehay TD. Monte Verde: A Late Pleistocene Settlement in Chile. Volume II: The Archaeological Context. Washington, DC: Smithsonian Institution Press; 1997.
|
| 39. |
Dillehay TD, Ocampo C, Saavedra J, Sawakuchi AO, Vega RM, Pino M, et al. New Archaeological Evidence for an Early Human Presence at Monte Verde, Chile. PLoS One. 2015;10(12):e0141923.
[Google Scholar]
[CrossRef]
|
| 40. |
Mena F, Reyes O. Montículos y cuevas funerarias en Patagonia: una visión desde cueva baño nuevo-1, XI región. Chungará (Arica). 2001;33(1):21-30.
[Google Scholar]
[CrossRef]
|
| 41. |
Reyes O, Méndez C, Mena F, Moraga M. The bioanthropological evidence of a ca. 10,000 CALYBP ten-individual group in central Patagonia. Southbound: late pleistocene peopling of latin America. 2012;167-171.
[Google Scholar]
|
| 42. |
Borrero LA. The Prehistoric Exploration and Colonization of Fuego-Patagonia. J World Prehist. 1999;13:321-355.
[Google Scholar]
[CrossRef]
|
| 43. |
Borrero LA. The Pleistocene—Holocene Transition in Southern South America. In: Straus LG, Eriksen BV, Erlandson JM, Yesner DR, editor. Humans at the End of the Ice Age: The Archaeology of the Pleistocene—Holocene Transition. Boston, MA: Springer US; 1996. p.339-354.
[Google Scholar]
[CrossRef]
|
| 44. |
Borrero LA, Zárate M, Miotti L, Massone M. The pleistocene–holocene transition and human occupations in the southern cone of South America. Quat Int. 1998;49-50:191-199.
[Google Scholar]
[CrossRef]
|
| 45. |
Miotti L. Piedra Museo locality: a special place in the New World. Current Research in the Pleistocene. 1995;12:37-40.
[Google Scholar]
|
| 46. |
Miotti L, Salemme M. Hunting and butchering events at late Pleistocene and early Holocene in Piedra Museo (Patagonia, southernmost south America). In: Bonnichsen R, editor. Paleoamerican Prehistory: colonization models, biological populations, and human adaptations. College Station, TX, USA: Center for the Study of the First Americans, University of Texas A&M; 2005. p.141-151.
[Google Scholar]
|
| 47. |
Paunero RS. The Cerro Tres Tetas locality (C3T) in the central Plateau of Santa Cruz, Argentina. Where the South Winds Blow: Ancient Evidence for Paleo South Americans. College Station, TX: Texas A&M University Press; 2004. p.133-141.
[Google Scholar]
|
| 48. |
Prieto A. Cazadores tempranos y tardíos en Cueva del Lago Sofía 1. Ans Inst Pat Ser Cs Soc [Internet]; 1991 [cited 2023 Nov 14]; Available from: https://cir.nii.ac.jp/crid/1573387448926178944.
|
| 49. |
Nami HG, Nakamura T. Cronología radiocarbónica con AMS sobre muestras de hueso procedentes del sitio Cueva del Medio (Ultima Esperanza, Chile). An Inst Patagonia. 1995;125:133.
[Google Scholar]
|
| 50. |
Waters MR, Amorosi T. Redating Fell’s Cave, Chile and the Chronological Placement of the Fishtail Projectile Point. Am Antiq. 2015;80(2):376-386.
[Google Scholar]
[CrossRef]
|
| 51. |
Massone M, Prieto A, Jackson D, Cárdenas G, Arroyo M, Cárdenas P. Los cazadores tempranos y sus fogatas: una nueva historia para la Cueva Tres Arroyos 1, Tierra del Fuego. Boletín de la Sociedad Chilena de Arqueología. 1998;26:11-18.
[Google Scholar]
|
| 52. |
Borrero LA. Taphonomy of the Tres Arroyos 1 Rockshelter, Tierra del Fuego, Chile. Quat Int. 2003;109-110:87-93.
[Google Scholar]
[CrossRef]
|
| 53. |
Massone MM. Vol. VII Los cazadores después del hielo. Museo de Historia Natural. Santiago: Centro de Investigaciones Diego Barros; 2004. 174 p.
[Google Scholar]
|
| 54. |
Rizzo F. Patrones de poblamiento del noroeste y centro-oeste de la Patagonia a partir del registro funerario y no funerario de cazadores-recolectores: Un modelo regional para los últimos 12.500 años. Latin American Antiquity. 2023;34(1):212-230.
[Google Scholar]
[CrossRef]
|
| 55. |
Méndez MC, Reyes BO, Nuevo Delaunay A, Trejo VV, Barberena R, Velásquez MH. Ocupaciones humanas en la margen occidental de Patagonia Central: eventos de poblamiento en alto río Cisnes. Magallania. 2011;39(2):223-242.
[Google Scholar]
[CrossRef]
|
| 56. |
Borrero LA, Nuevo Delaunay A, Méndez C. Ethnographical and historical accounts for understanding the exploration of new lands: The case of Central Western Patagonia, Southernmost South America. J Anthropol Archaeol. 2019;54:1-16.
[Google Scholar]
[CrossRef]
|
| 57. |
Whitlock C, Bianchi MM, Bartlein PJ, Markgraf V, Marlon J, Walsh M, et al. Postglacial vegetation, climate, and fire history along the east side of the Andes (lat 41–42.5°S), Argentina. Quat Res. 2006;66(2):187-201.
[Google Scholar]
[CrossRef]
|
| 58. |
Belardi JB. Cuevas, aleros, distribuciones y poblamiento. In: Gómez Otero J, editor. Arqueología Sólo Patagonia. Puerto Madryn: Centro Nacional Patagónico; 1996. p.43-48.
[Google Scholar]
|
| 59. |
Fiore D. Poblamiento de imágenes: arte rupestre y colonización de la Patagonia. Variabilidad y ritmos de cambio en tiempo y espacio. Tramas en la piedra Producción y usos. 2006;43-62.
[Google Scholar]
|
| 60. |
Powell JF, Neves WA. Craniofacial morphology of the first Americans: pattern and process in the peopling of the New World. Am J Phys Anthropol. 1999;29:153-158.
[Google Scholar]
|
| 61. |
Neves WA, Pucciarelli HM. Morphological affinities of the first Americans: an exploratory analysis based on early South American human remains. J Hum Evol. 1991;21(4):261-273.
[Google Scholar]
[CrossRef]
|
| 62. |
Neves WA, Prous A, González-José R, Kipnis R, Powell J. Early Holocene human skeletal remains from Santana do Riacho, Brazil: implications for the settlement of the New World. J Hum Evol. 2003;45(1):19-42.
[Google Scholar]
[CrossRef]
|
| 63. |
Lalueza C., Pérez-Pérez A, Prats E, Cornudella L, Turbon D. Lack of founding Amerindian mitochondrial DNA lineages in extinct Aborigines from Tierra del Fuego–Patagonia. Hum Mol Genet. 1997;6(1):41-46.
[Google Scholar]
[CrossRef]
|
| 64. |
González-José R, Bortolini MC, Santos FR, Bonatto SL. The peopling of America: craniofacial shape variation on a continental scale and its interpretation from an interdisciplinary view. Am J Phys Anthropol. 2008;137(2):175-187.
[Google Scholar]
[CrossRef]
|
| 65. |
Perez SI, Bernal V, Gonzalez PN, Sardi M, Politis GG. Discrepancy between cranial and DNA data of early Americans: implications for American peopling. PLoS One. 2009;4(5):e5746.
[Google Scholar]
[CrossRef]
|
| 66. |
Marangoni A, Caramelli D, Manzi G. Homo sapiens in the Americas. Overview of the earliest human expansion in the New World J Anthropol Sci. 2014;92:79-97.
[Google Scholar]
|
| 67. |
Politis GG, Prates L, Perez SI. El poblamiento de América: arqueología y bio-antropología de los primeros americanos. Buenos Aires: Eudeba; 2009.
|
| 68. |
Moraga ML, Rocco P, Miquel JF, Nervi F, Llop E, Chakraborty R, et al. Mitochondrial DNA polymorphisms in Chilean aboriginal populations: implications for the peopling of the southern cone of the continent. Am J Phys Anthropol. 2000;113(1):19-29.
[Google Scholar]
|
| 69. |
Raghavan M, Steinrücken M, Harris K, Schiffels S, Rasmussen S, DeGiorgio M, et al. POPULATION GENETICS. Genomic evidence for the Pleistocene and recent population history of Native Americans. Science. 2015;349(6250):aab3884.
[Google Scholar]
|
| 70. |
Moreno-Mayar JV, Vinner L, de Barros Damgaard P, de la Fuente C, Chan J, Spence JP, et al. Early human dispersals within the Americas. Science [Internet]. 2018 Dec 7;362(6419). [cited 2023 Nov 14]. Available from: http://dx.doi.org/10.1126/science.aav2621.
[Google Scholar]
|
| 71. |
Posth C, Nakatsuka N, Lazaridis I, Skoglund P, Mallick S, Lamnidis TC, et al. Reconstructing the Deep Population History of Central and South America. Cell. 2018;175(5):1185-1197.e22.
[Google Scholar]
|
| 72. |
García-Bour J, Pérez-Pérez A, Alvarez S, Fernández E, López-Parra AM, Arroyo-Pardo E, et al. Early population differentiation in extinct aborigines from Tierra del Fuego-Patagonia: ancient mtDNA sequences and Y-chromosome STR characterization. Am J Phys Anthropol. 2004;123(4):361-370.
[Google Scholar]
[CrossRef]
|
| 73. |
de la Fuente C, Galimany J, Kemp BM, Judd K, Reyes O, Moraga M. Ancient marine hunter-gatherers from Patagonia and Tierra Del Fuego: Diversity and differentiation using uniparentally inherited genetic markers. Am J Phys Anthropol. 2015;158(4):719-729.
[Google Scholar]
[CrossRef]
|
| 74. |
Balentine CM, Alfonso-Durruty M, Reynolds AW, Vilar M, Morello F, Román MS, et al. Evaluating population histories in Patagonia and Tierra del Fuego, Chile, using ancient mitochondrial and Y-chromosomal DNA. Am J Biol Anthropol. 2022;180(1):144-161.
[Google Scholar]
[CrossRef]
|
| 75. |
de la Fuente C, Ávila-Arcos MC, Galimany J, Carpenter ML, Homburger JR, Blanco A, et al. Genomic insights into the origin and diversification of late maritime hunter-gatherers from the Chilean Patagonia. Proc Natl Acad Sci USA. 2018;115(17):E4006-E4012.
[Google Scholar]
[CrossRef]
|
| 76. |
Nakatsuka N, Luisi P, Motti JMB, Salemme M, Santiago F, D’Angelo Del Campo MD, et al. Ancient genomes in South Patagonia reveal population movements associated with technological shifts and geography. Nat Commun. 2020;11:3868.
[Google Scholar]
[CrossRef]
|
| 77. |
Paz Sepúlveda PB, Mayordomo AC, Sala C, Sosa EJ, Zaiat JJ, Cuello M, et al. Human Y chromosome sequences from Q Haplogroup reveal a South American settlement pre-18,000 years ago and a profound genomic impact during the Younger Dryas. PLoS One. 2022;17(8):e0271971.
[Google Scholar]
[CrossRef]
|
| 78. |
Moraga ML, de la Fuente C, Galimany J, Orellana-Soto M, Reyes O. The ancient inhabitants of the Chonos Archipelago (~43°–47°S): Characterization of maritime hunter-gatherers from Western Patagonia using mitochondrial DNA. Am J Biol Anthropol. 2022;180(2):286-297.
[Google Scholar]
[CrossRef]
|
| 79. |
de Saint Pierre M, Gandini F, Perego UA, Bodner M, Gómez-Carballa A, Corach D, et al. Arrival of Paleo-Indians to the southern cone of South America: new clues from mitogenomes. PLoS One. 2012;7(12):e51311.
[Google Scholar]
[CrossRef]
|
| 80. |
de Saint Pierre M, Bravi CM, Motti JMB, Fuku N, Tanaka M, Llop E, et al. An alternative model for the early peopling of southern South America revealed by analyses of three mitochondrial DNA haplogroups. PLoS One. 2012;7(9):e43486.
[Google Scholar]
[CrossRef]
|
| 81. |
Arencibia V, Crespo CM, García Guraieb S, Russo MG, Dejean CB, Goñi R. Análisis genético poblacional de grupos cazadores recolectores del Holoceno tardío del Lago Salitroso (Santa Cruz, Argentina). Revista Argentina de Antropología Biológica. 2019;21(2):004.
[Google Scholar]
[CrossRef]
|
| 82. |
Arencibia V, Muñoz M, Crespo CM, Russo MG, Vera P, Lia VV, et al. Novel B2 mitogenomes from Continental southern Patagonia’s Late Holocene: New insights into the peopling of the Southern Cone. Am J Biol Anthropol. 2023.
[Google Scholar]
[CrossRef]
|
| 83. |
Parolin ML, Lorenzo RIF, Bravi CM, Mendez FL, Dahinten SL, Gómez-Otero J, Bustamante C, et al. Ancient mitogenomes of Argentine Patagonia (6070-310 YBP) reveal the early contribution of lineages not previously found in South America. PeerJ Preprints. 2017;5:e3098v1.
[Google Scholar]
[CrossRef]
|
| 84. |
Franco NV, Galimany J, Moraga M, Borrero LA. La integración de información cultural y genética en el sur de la cuenca superior del río Santa Cruz: primeros resultados. Décimas Jornadas de Arqueología de la Patagonia. Puerto Madryn: IDEAUS-CONICET; 2017.
[Google Scholar]
|
| 85. |
Perego UA, Achilli A, Angerhofer N, Accetturo M, Pala M, Olivieri A, et al. Distinctive Paleo-Indian migration routes from Beringia marked by two rare mtDNA haplogroups. Curr Biol. 2009;19(1):1-8.
[Google Scholar]
[CrossRef]
|
| 86. |
Motti JMB, Hagelberg E, Lindo J, Malhi RS, Bravi CM, Guichón RA. First complete mitochondrial genome sequence from human skeletal remains of the Coast of Santa Cruz, Argentina. Magallania. 2015;43(2):119-131.
[Google Scholar]
[CrossRef]
|
| 87. |
Rothhammer F, Dillehay TD. The late Pleistocene colonization of South America: an interdisciplinary perspective. Ann Hum Genet. 2009;73(5):540-549.
[Google Scholar]
[CrossRef]
|
| 88. |
Miotti L, Salemme MC. When Patagonia was colonized: people mobility at high latitudes during Pleistocene/Holocene transition. Quat Int. 2003;109-110:95-111.
[Google Scholar]
[CrossRef]
|
| 89. |
Rull V. Late pleistocene and Holocene climates of Venezuela. Quat Int. 1996;31:85-94.
[Google Scholar]
[CrossRef]
|
| 90. |
Prieto AR. Late Quaternary Vegetational and Climatic Changes in the Pampa Grassland of Argentina. Quat Res. 1996;45(1):73-88.
[Google Scholar]
[CrossRef]
|
| 91. |
Latrubesse EM, Ramonell CG. A climatic model for southwestern Amazonia in Last Glacial times. Quat Int. 1994;21:163-169.
[Google Scholar]
[CrossRef]
|
| 92. |
Weng C, Bush MB, Silman MR. An analysis of modern pollen rain on an elevational gradient in southern Peru. J Trop Ecol. 2004;20(01):113-124.
[Google Scholar]
[CrossRef]
|
| 93. |
Bush MB, Flenley J. Tropical Rainforest Responses to Climatic Change. Berlin, Heidelberg: Springer; 2007. 396 p.
[CrossRef]
|
| 94. |
Rasmussen M, Anzick SL, Waters MR, Skoglund P, DeGiorgio M, Stafford TW Jr, et al. The genome of a Late Pleistocene human from a Clovis burial site in western Montana. Nature. 2014;506(7487):225-229.
[Google Scholar]
[CrossRef]
|
| 95. |
Arango-Isaza E, Capodiferro MR, Aninao MJ, Babiker H, Aeschbacher S, Achilli A, et al. The genetic history of the Southern Andes from present-day Mapuche ancestry. Curr Biol. 2023;33(13):2602-2615.e5.
[Google Scholar]
[CrossRef]
|
| 96. |
Capodiferro MR, Aram B, Raveane A, Rambaldi Migliore N, Colombo G, Ongaro L, et al. Archaeogenomic distinctiveness of the Isthmo-Colombian area. Cell. 2021;184(7):1706-1723.e24.
[Google Scholar]
[CrossRef]
|
| 97. |
Ferraz T, Suarez Villagran X, Nägele K, Radzevičiūtė R, Barbosa Lemes R, Salazar-García DC, et al. Genomic history of coastal societies from eastern South America. Nat Ecol Evol. 2023;7(8):1315-1330.
[Google Scholar]
[CrossRef]
|
| 98. |
Willerslev E, Meltzer DJ. Peopling of the Americas as inferred from ancient genomics. Nature. 2021;594(7863):356-364.
[Google Scholar]
[CrossRef]
|
| 99. |
Moraga M, De Saint Pierre M, Torres F, Ríos J. Vínculos de parentesco por vía materna entre los últimos descendientes de la etnia Kawésqar y algunos entierros en los canales patagónicos: evidencia desde el estudio de linajes mitocondriales. Magallania (Punta Arenas). 2010;38(2):103-114.
[Google Scholar]
[CrossRef]
|
| 100. |
Crespo CM, Lanata JL, Cardozo DG, Avena SA, Dejean CB. Ancient maternal lineages in hunter-gatherer groups of Argentinean Patagonia. Settlement, population continuity and divergence. J Archaeol Sci Rep. 2018;18:689-695.
[Google Scholar]
[CrossRef]
|
| 101. |
Motti JMB, Winingear S, Valenzuela LO, Nieves-Colón MA, Harkins KM, García Laborde P, et al. Identification of the geographic origins of people buried in the cemetery of the Salesian Mission of Tierra del Fuego through the analyses of mtDNA and stable isotopes. J Archaeol Sci Rep. 2020;33:102559.
[Google Scholar]
[CrossRef]
|
| 102. |
Crespo CM, Russo MG, Hajduk A, Lanata JL, Dejean CB. Variabilidad mitocondrial en muestras pre-colombinas de la Patagonia Argentina: Hacia una visión de su poblamiento desde el ADN antiguo. Revista argentina de antropología biológica. 2016;19(1).
[Google Scholar]
[CrossRef]
|
| 103. |
Motti J, Muñoz S, Cruz I, D’Angelo del Campo M, Borrero L, Bravi C, et al. Análisis de ADN mitocondrial en restos humanos del Holoceno Tardío del sur de Santa Cruz. In: Gómez Otero J, Svoboda A, Banegas A, editor. Arqueología de la Patagonia: el pasado en las arenas. Puerto Madryn: Instituto de Diversidad y Evolución Austral; 2019. p.493-503.
[Google Scholar]
|
| 104. |
Parolin ML, Galimany J, Gómez Otero J, Dahinten S, Millán AG, Moraga M. Primeras secuencias mitocondriales de la región control completa en muestras humanas del Holoceno tardío de la costa norte y centro de Patagonia, Argentina. In: Gómez Otero J, Svoboda A, Banegas A, editor. Arqueología de la Patagonia: El pasado en las arenas. Puerto Madryn: Instituto de Diversidad y Evolución Austral; 2019. p.469-480.
[Google Scholar]
|
| 105. |
Clapperton CM. La última glaciación y deglaciación en el Estrecho de Magallanes: Implicaciones para el poblamiento de Tierra del Fuego. Anales del Instituto de la Patagonia [Internet]; 1992 [cited 2023 Nov 14]. Available from: http://www.bibliotecadigital.umag.cl/bitstream/handle/20.500.11893/1002/Clapperton_Anales_1992_vol21_pp113-128.pdf?sequence=1&isAllowed=y.
|
| 106. |
Porter SC, Stuiver M, Heusser CJ. Holocene sea-level changes along the Strait of Magellan and Beagle Channel, southernmost South America. Quat Res. 1984;22(1):59-67.
[Google Scholar]
[CrossRef]
|
| 107. |
Crespo CM, Cardozo DG, Tessone A, Vázquez M, Kisielinski C, Arencibia V, et al. Distribution of maternal lineages in hunter-gatherer societies of the southern coast of Tierra del Fuego, Argentina. Am J Phys Anthropol. 2020;173(4):709-720.
[Google Scholar]
[CrossRef]
|
| 108. |
Tamburrini C, de Saint Pierre M, Bravi CM, Bailliet G, Jurado Medina L, Velázquez IF, et al. Uniparental origins of the admixed Argentine Patagonia. Am J Hum Biol. 2022;34(4):e23682.
[Google Scholar]
[CrossRef]
|
| 109. |
Avena SA, Parolin ML, Dejean CB, Ríos Part MC, Fabrikant G, Goicoechea AS, et al. Mezcla génica y linajes uniparentales en Comodoro Rivadavia (provincia de Chubut, Argentina). Rev Argent Endocrinol Metab [Internet]; 2009 [cited 2023 Nov 14]; Available from: https://www.redalyc.org/pdf/3822/382239053004.pdf.
|
| 110. |
Parolin ML, Avena SA, Fleischer S, Pretell M, Rocca FDF, Rodríguez DA, et al. Análisis de la diversidad biológica y mestizaje en la ciudad de Puerto Madryn (Prov. de Chubut, Argentina). Revista argentina de antropología biológica. 2013;15(1):61-75.
[Google Scholar]
|
| 111. |
Toscanini U, Brisighelli F, Moreno F, Pantoja-Astudillo JA, Morales EA, Bustos P, et al. Analysis of Y-chromosome STRs in Chile confirms an extensive introgression of European male lineages in urban populations. Forensic Sci Int Genet. 2016;21:76-80.
[Google Scholar]
[CrossRef]
|
| 112. |
Rodrigues P, Velázquez IF, Ribeiro J, Simão F, Amorim A, Carvalho EF, et al. Tierra Del Fuego: What Is Left from the Precolonial Male Lineages? Genes. 2022;13(10):1712.
[Google Scholar]
[CrossRef]
|
| 113. |
Ramallo V, Mucci JM, García A, Muzzio M, Motti JMB, Santos MR, et al. Comparison of Y-chromosome haplogroup frequencies in eight Provinces of Argentina. Forensic Sci Int Genet Suppl Ser. 2009;2(1):431-432.
[Google Scholar]
[CrossRef]
|
| 114. |
Di Fabio Rocca F, Albeza MV, Postillone MB, Acreche N, Lafage L, Parolín ML, et al. Historia poblacional y análisis antropogenético de la ciudad de Salta. Andes. 2016.
[Google Scholar]
|
| 115. |
Cabana GS, Merriwether DA, Hunley K, Demarchi DA. Is the genetic structure of Gran Chaco populations unique? Interregional perspectives on native South American mitochondrial DNA variation. Am J Phys Anthropol. 2006;131(1):108-119.
[Google Scholar]
[CrossRef]
|
| 116. |
Sevini F, Yao DY, Lomartire L, Barbieri A, Vianello D, Ferri G, et al. Analysis of population substructure in two sympatric populations of Gran Chaco, Argentina. PLoS One. 2013;8(5):e64054.
[Google Scholar]
[CrossRef]
|
| 117. |
Motti JMB, Rodenak B, Muzzio M, Ramallo V, Santos MR, Castro C, et al. The genetic composition of Argentina prior to the massive immigration era: Insights from matrilineages of extant criollos in central-western Argentina. Forensic Sci Int Genet Suppl Ser. 2009;2(1):342-343.
[Google Scholar]
[CrossRef]
|
| 118. |
Catelli ML, Alvarez-Iglesias V, Gómez-Carballa A, Mosquera-Miguel A, Romanini C, Borosky A, et al. The impact of modern migrations on present-day multi-ethnic Argentina as recorded on the mitochondrial DNA genome. BMC Genet. 2011;12(1):77.
[Google Scholar]
[CrossRef]
|
| 119. |
Pauro M, García A, Bravi CM, Demarchi D. Distribución de haplogrupos mitocondriales alóctonos en poblaciones rurales de Córdoba y San Luis/Distribution of introduced mitochondrial haplogroups in rural populations of Córdoba and San Luis. Rev Argent Antropol Biol. 2010;12(1):47-55.
[Google Scholar]
|
| 120. |
Motti J, Muzzio M, Ramallo V, Rodenak Kladniew B, Alfaro EL, Dipierri JE, et al. Origen y distribución espacial de linajes maternos nativos en el noroeste y centro oeste argentinos. Revista Argentina De Antropología Biológica. 2012;15(1):3-14.
[Google Scholar]
|
| 121. |
Lalueza Fox C. Mitochondrial DNA haplogroups in four tribes from Tierra del Fuego—Patagonia: Inferences about the peopling of the Americas. Hum Biol. 1996;11:217-224.
[Google Scholar]
[CrossRef]
|
| 122. |
Reich D, Patterson N, Campbell D, Tandon A, Mazieres S, Ray N, et al. Reconstructing Native American population history. Nature. 2012;488:370-374.
[Google Scholar]
[CrossRef]
|
| 123. |
Di Fabio Rocca F, Russo MG, Arencibia V, Seldes V. Ancient DNA studies: Use of ethnonyms and collaborative research in South America. Anthropol. Archaeol. 2021;61:101265.
[Google Scholar]
[CrossRef]
|
| 124. |
Arencibia V, Muñoz Hidalgo M, Crespo C, Maldonado L, Lichtenstein G, Kamenetzky L, et al. Ancient Human Mitochondrial Genome of the Beagle Channel (Tierra Del Fuego, Argentina): Enrichment, Sequencing And Analysis, a Collaborative Project. Lat Am Antiq. 2023;1-14.
[Google Scholar]
[CrossRef]
|
| 125. |
Luisi P, García A, Berros JM, Motti JMB, Demarchi DA, Alfaro E, et al. Fine-scale genomic analyses of admixed individuals reveal unrecognized genetic ancestry components in Argentina. PLoS One. 2020;15(7):e0233808.
[Google Scholar]
[CrossRef]
|
| 126. |
Gigoux C. “Condemned to Disappear”: Indigenous Genocide in Tierra del Fuego. J Genocide Res. 2022;24(1):1-22.
[Google Scholar]
[CrossRef]
|
| 127. |
Tamburrini C, Dahinten SL, Saihueque RRR, Ávila-Arcos MC, Parolin ML. Towards an ethical and legal framework in archeogenomics: A local case in the Atlantic coast of central Patagonia. Am J Biol Anthropol. 2023;182(2):161-176.
[Google Scholar]
[CrossRef]
|
| 128. |
Endere ML, Ayala P. Normativa legal, recaudos éticos y práctica arqueológica: un estudio comparativo de Argentina y Chile. Chungará (Arica). 2012;44(1):39-57.
[Google Scholar]
[CrossRef]
|
| 129. |
Eisenmann S, Bánffy E, van Dommelen P, Hofmann KP, Maran J, Lazaridis I, et al. Reconciling material cultures in archaeology with genetic data: The nomenclature of clusters emerging from archaeogenomic analysis. Sci Rep. 2018;8(1):13003.
[Google Scholar]
[CrossRef]
|
| 130. |
Gopalakrishnan S, Ebenesersdóttir SS, Lundstrøm IKC, Turner-Walker G, Moore KHS, Luisi P, et al. The population genomic legacy of the second plague pandemic. Curr Biol. 2022;32(21):4743-4751.e6.
[Google Scholar]
[CrossRef]
|
| 131. |
Fu Q, Hajdinjak M, Moldovan OT, Constantin S, Mallick S, Skoglund P, et al. An early modern human from Romania with a recent Neanderthal ancestor. Nature. 2015;524(7564):216-219.
[Google Scholar]
[CrossRef]
|
| 132. |
Haak W, Lazaridis I, Patterson N, Rohland N, Mallick S, Llamas B, et al. Massive migration from the steppe was a source for Indo-European languages in Europe. Nature. 2015;522(7555):207-211.
[Google Scholar]
[CrossRef]
|
| 133. |
Mathieson I, Lazaridis I, Rohland N, Mallick S, Patterson N, Roodenberg SA, et al. Genome-wide patterns of selection in 230 ancient Eurasians. Nature. 2015;528(7583):499-503.
[Google Scholar]
[CrossRef]
|
| 134. |
Pickrell JK, Reich D. Toward a new history and geography of human genes informed by ancient DNA. Trends Genet. 2014;30(9):377-389.
[Google Scholar]
[CrossRef]
|
| 135. |
Ávila-Arcos MC, Raghavan M, Schlebusch C. Going local with ancient DNA: A review of human histories from regional perspectives. Science. 2023;382(6666):53-58.
[Google Scholar]
[CrossRef]
|
| 136. |
Günther T, Nettelblad C. The presence and impact of reference bias on population genomic studies of prehistoric human populations. PLoS Genet. 2019;15(7):e1008302.
[Google Scholar]
[CrossRef]
|
| 137. |
Davidson R, Williams MP, Roca-Rada X, Kassadjikova K, Tobler R, Fehren-Schmitz L, Llamas B. Allelic bias when performing in-solution enrichment of ancient human DNA. Mol Ecol Resour. 2023;23:1823-1840.
[Google Scholar]
[CrossRef]
|
| 138. |
Lindo J, Huerta-Sánchez E, Nakagome S, Rasmussen M, Petzelt B, Mitchell J, et al. A time transect of exomes from a Native American population before and after European contact. Nat Commun. 2016;7(1):13175.
[Google Scholar]
[CrossRef]
|
| 139. |
Ringbauer H, Huang Y, Akbari A, Mallick S, Patterson N, Reich D. ancIBD - Screening for identity by descent segments in human ancient DNA. bioRxiv [Internet]; 2023 [cited 2023 Nov 14]; Available from: http://dx.doi.org/10.1101/2023.03.08.531671.
[CrossRef]
|
| 140. |
Racimo F, Sikora M, Vander Linden M, Schroeder H, Lalueza-Fox C. Beyond broad strokes: sociocultural insights from the study of ancient genomes. Nat Rev Genet. 2020;21(6):355-366.
[Google Scholar]
[CrossRef]
|
| 141. |
Monroy Kuhn JM, Jakobsson M, Günther T. Estimating genetic kin relationships in prehistoric populations. PLoS One. 2018;13(4):e0195491.
[Google Scholar]
[CrossRef]
|
| 142. |
Popli D, Peyrégne S, Peter BM. KIN: a method to infer relatedness from low-coverage ancient DNA. Genome Biol. 2023;24(1):10.
[Google Scholar]
|
| 143. |
Lipatov M, Sanjeev K, Patro R, Veeramah KR. Maximum Likelihood Estimation of Biological Relatedness from Low Coverage Sequencing Data. bioRxiv. 2015.
[Google Scholar]
[CrossRef]
|
| 144. |
Fernandes DM, Cheronet O, Gelabert P, Pinhasi R. TKGWV2: an ancient DNA relatedness pipeline for ultra-low coverage whole genome shotgun data. Sci Rep. 2021;11(1):21262.
[Google Scholar]
[CrossRef]
|
| 145. |
Waples RK, Albrechtsen A, Moltke I. Allele frequency-free inference of close familial relationships from genotypes or low-depth sequencing data. Mol Ecol. 2019;28(1):35-48.
[Google Scholar]
[CrossRef]
|
| 146. |
Kerner G, Choin J, Quintana-Murci L. Ancient DNA as a tool for medical research. Nat Med. 2023;29(5):1048-1051.
[Google Scholar]
[CrossRef]
|
| 147. |
Leavitt PA, Covarrubias R, Perez YA, Fryberg SA. “frozen in time”: The impact of native American media representations on identity and self-understanding. J Soc Issues. 2015;71(1):39-53.
[Google Scholar]
[CrossRef]
|
| 148. |
Varela HH, Cocilovo JA, Guichon RA. Evaluación de la información somatométrica por Gusinde sobre los aborígenes de Tierra del Fuego. Anales del Instituto de la Patagonia. 1993;22:193-205.
[Google Scholar]
|
| 149. |
Polderman TJC, Benyamin B, de Leeuw CA, Sullivan PF, van Bochoven A, Visscher PM, et al. Meta-analysis of the heritability of human traits based on fifty years of twin studies. Nat Genet. 2015;47(7):702-709.
[Google Scholar]
[CrossRef]
|
| 150. |
Watanabe K, Stringer S, Frei O, Umićević Mirkov M, de Leeuw C, Polderman TJC, et al. A global overview of pleiotropy and genetic architecture in complex traits. Nat Genet. 2019;51(9):1339-1348.
[Google Scholar]
[CrossRef]
|
| 151. |
Turchin MC, Chiang CWK, Palmer CD, Sankararaman S, Reich D, Genetic Investigation of ANthropometric Traits (GIANT) Consortium, et al. Evidence of widespread selection on standing variation in Europe at height-associated SNPs. Nat Genet. 2012;44(9):1015-1019.
[Google Scholar]
[CrossRef]
|
| 152. |
Sohail M, Maier RM, Ganna A, Bloemendal A, Martin AR, Turchin MC, et al. Polygenic adaptation on height is overestimated due to uncorrected stratification in genome-wide association studies. eLife. 2019;8:e39702.
[Google Scholar]
[CrossRef]
|
| 153. |
Bergmann C. Über die Verhältnisse der Wärmeökonomie der Thiere zu ihrer Größe. Göttingen: Vandenhoeck und Ruprecht; 1848. 116 p.
|
| 154. |
Béguelin M, Barrientos G. Variación morfométrica postcraneal en muestras tardías de restos humanos de Patagonia: una aproximación biogeográfica. Intersecc Antropol. 2006;49-62.
[Google Scholar]
|
| 155. |
Alfonso-Durruty M, Giles BT, San Román M, Morello F. Assessment of an east-west phenotypic variation in body height, body form and body mass among prehistoric hunter-gatherers of Patagonia and Tierra del Fuego, Chile. Chungara, Revista de Antropología Chilena. 2017;49(4):1-12.
[Google Scholar]
[CrossRef]
|
| 156. |
García-Martínez D, Nalla S, Ferreira MT, Guichón RA, D’Angelo Del Campo MD, Bastir M. Eco-geographic adaptations in the human ribcage throughout a 3D geometric morphometric approach. Am J Phys Anthropol. 2018;166(2):323-336.
[Google Scholar]
[CrossRef]
|
| 157. |
Zaidi AA, Mattern BC, Claes P, McEvoy B, Hughes C, Shriver MD. Correction: Investigating the case of human nose shape and climate adaptation. PLoS Genet. 2018;14(1):e1007207.
[Google Scholar]
[CrossRef]
|
| 158. |
Li Q, Chen J, Faux P, Delgado ME, Bonfante B, Fuentes-Guajardo M, et al. Automatic landmarking identifies new loci associated with face morphology and implicates Neanderthal introgression in human nasal shape. Commun Biol. 2023 May 8;6(1):481.
[Google Scholar]
[CrossRef]
|
| 159. |
Mallick S, Micco A, Mah M, Ringbauer H, Lazaridis I, Olalde I, et al. The Allen Ancient DNA Resource (AADR): A curated compendium of ancient human genomes. bioRxiv. 2023.
[Google Scholar]
[CrossRef]
|
| 160. |
Chang CC, Chow CC, Tellier LC, Vattikuti S, Purcell SM, Lee JJ. Second-generation PLINK: rising to the challenge of larger and richer datasets. Gigascience. 2015;4(1):7.
[Google Scholar]
[CrossRef]
|


 ,
Josefina Motti
2,3,†
,
Josefina Motti
2,3,†