
Human Population Genetics and Genomics ISSN 2770-5005
Human Population Genetics and Genomics 2024;4(4):0010 | https://doi.org/10.47248/hpgg2404040010
Opinion Open Access
Interpreting the demic diffusion of early farming in Europe with a three-population model
Kenichi Aoki

Academic Editor(s): Alan Templeton
Received: May 6, 2024 | Accepted: Oct 2, 2024 | Published: Oct 8, 2024
This article belongs to the Special Issue Luca Cavalli-Sforza’s legacy, 100 years after his birth
© 2024 by the author(s). This is an Open Access article distributed under the terms of the Creative Commons License Attribution 4.0 International (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium or format, provided the original work is correctly credited.
Cite this article: Aoki K. Interpreting the demic diffusion of early farming in Europe with a three-population model. Hum Popul Genet Genom 2024; 4(4):0010. https://doi.org/10.47248/hpgg2404040010
In 1971, Ammerman and Cavalli-Sforza demonstrated that reaction-diffusion equations could be usefully applied to the archaeological question of the spread of early farming in Europe. Their basic premise was demic diffusion, i.e., the iterative short-range colonization of virgin land by the descendants of the original Near Eastern farmers. This hypothesis has been vindicated by ancient DNA studies, which show limited acculturation of the autochthonous hunter-gatherers, who when converted to farming were apparently assimilated into preexisting farming communities. In this brief report, I describe a reaction-diffusion model incorporating various interactions among the Near Eastern farmers, converted farmers, and hunter-gatherers. Predictions, derived in terms of the model parameters, are examined vis-à-vis the ancient DNA and archaeological evidence. Of particular interest is the theoretical requirement that the hunter-gatherers behaved more competitively toward the converted farmers than the Near Eastern (specifically Anatolian) famers.
Based on “Ammerman AJ, Cavalli-Sforza LL. Measuring the rate of spread of early farming in Europe. Man 1971; 6: 674-688.”
Keywordsdemic diffusion, ancient DNA, reaction-diffusion model, model predictions
Farming, based on domesticated plants and animals, was introduced into Europe from the Near East perhaps nine thousand years ago and spread throughout much of the continent over a period of several thousand years. Who were the bearers of the farming culture and what was the fate of the indigenous hunter-gatherers? There is now agreement on the main issues (see below), and our current understanding owes much to the impressive application of the inductive-deductive approach by the archaeologist Albert Jay Ammerman and the geneticist Luigi Luca Cavalli-Sforza beginning in 1971.
The geographical spread of a novel cultural activity such as farming can be explained in at least three contrasting (but not necessarily mutually exclusive) ways, which Ammerman and Cavalli-Sforza [1] called ‘cultural diffusion’, ‘demic diffusion’, and ‘colonization’. Cultural diffusion entails the transmission of know-how from a farming to a neighboring non-farming community without significant population displacement. Demic diffusion occurs when a farming community undergoes population growth and a minority of its members move to a random adjacent area, carrying their know-how with them. In both cases, the process is an iterative one. Finally, colonization connotes a one-time long-distance migration by the entire community or a cohesive sub-section thereof. See Ammerman [2] for a recent in-depth discussion of mobility patterns.
Ammerman and Cavalli-Sforza [1] combined (i) an analysis of the spatial-temporal distribution of radiocarbon dates that record the initial spread of early farming in Europe (ii) with a mathematical model [3,4] that could explain the statistically significant rate constancy. Applying this model—the argument is described in the following three paragraphs—they suggested that early farming may have spread by demic diffusion. Their estimate of the speed was ~1 kilometer per year with a point of origin in the Levant.
In a sequel paper, Ammerman and Cavalli-Sforza [5] invoked the Fisher [6] equation,
Here n(x,t) represents the population density (of farmers) at location x and time t in an infinite one-dimensional space. The left-hand side of Equation (1) is the rate of change with time of this density (subscript t indicates the partial derivative with respect to time). The first term on the right-hand side gives the effect of random bidirectional dispersal (subscript xx indicates the second partial derivative of spatial location), where D is the diffusion coefficient (see below). The second term expresses logistic growth, where r denotes the intrinsic (without competition) growth rate, and K is the carrying capacity (density sustainable with the available technology) of the environment. The Fisher [6] model is a more realistic representation of population dynamics than the Kendall [3] and Skellam [4] models that lack the term n/K.
Numerical analysis of Equation (1) yields a wave with an advancing front that eventually attains a constant speed. Fisher [6] used a heuristic argument to show that the (minimum such) speed would be
which is effectively the speed observed in numerical work. Unfortunately, an explicit solution of Equation (1) is not available (for this speed). Ammerman and Cavalli-Sforza [5] provide cautious estimates of the two parameters, r and D, that appear in Equation (2).
Unmitigated application of the Fisher [6] model would entail that the spread of early farming in Europe occurred by the range expansion of the Near Eastern farmers, with little or no interaction with the indigenous hunter-gatherers, i.e., solely by demic diffusion. In the same papers [1,5], they discuss the possibility that the Near Eastern farmers admixed with the local Mesolithic groups that they encountered during their migratory advance. The latter scenario would have resulted in a progressive dilution of Near Eastern farmer ancestry and perhaps produced gene frequency clines detectable in modern Europeans.
Sgaramella-Zonta and Cavalli-Sforza [7] and Rendine et al. [8] addressed this issue with a two-population simulation model of farmers and hunter-gatherers. In their models, space is represented by cells arranged in a two-dimensional lattice. Migration occurs between neighboring cells. Within each cell, hunter-gatherers and farmers undergo logistic growth with subsistence-dependent parameters, and unidirectional conversion (acculturation) of hunter-gatherers to farming occurs at a certain rate. Gene frequency changes can occur among hunter-gatherers and farmers by the random sampling of diallelic genes at birth or in association with conversion. It is shown that the ratio of the carrying capacities of the two populations and the acculturation rate significantly affect the formation of gene frequency clines.
People were presumably not continuously distributed in space as assumed in a reaction-diffusion model such as Equation (1), but were clustered in settlements. Archaeological sites are the remains of such settlements. If we picture discrete settlements separated by equal distances, then the diffusion coefficient, D, corresponds to one-half and one-quarter of the variance in migration rate in one-dimensional and two-dimensional space, respectively.
In the models of Sgaramella-Zonta and Cavalli-Sforza [7] and of Rendine et al. [8], the population of farmers within a cell may comprise individuals of Near Eastern farmer ancestry and indigenous hunter-gatherer ancestry (and their admixed descendants). This was our motivation for proposing a three-population model [9,10], described below, which distinguishes the two kinds of farmers, and which in some respects makes the book-keeping easier. However, our model suffers from numerous inadequacies, numbered as limitations #1–#6, which will be addressed in turn as we come to them.
Pinhasi et al. [11] have collated more than 700 radiocarbon dates (calibrated, abbreviated to ‘cal’, and uncalibrated) of Neolithic sites in the Near East (including Anatolia), continental Europe, and the British Isles. In addition to the great-circle distances used by Ammerman and Cavalli-Sforza [1], they considered shortest-path distances that allow for short sea travel. When the latter are used with their vastly extended dataset, they find that the most likely point of origin for the introduction of early farming into Europe lies in a region that includes southeast Anatolia. This result is consistent with ancient DNA studies, and their estimate of the average rate of spread is 0.6–1.3 kilometers per year.
Early farming is believed to have been brought into Europe by farmers of Anatolian ancestry via the Aegean shores and the Balkans. According to Allentoft et al. [12], the earliest evidence for Anatolian-related ancestry in Europe, dated to 6700 BCE (Before Common Era), occurs at the site of Lepenski Vir in present-day Serbia. Subsequent spread within continental Europe initially followed two land routes, one along the Danube River associated with the Linearbandkeramik (LBK) culture, and the other along the northern coast of the Mediterranean Sea associated with Cardial (also called Impresso or Impressa) pottery, e.g., [13]. Along the Danubian route, farming had reached eastern France by ~5200 BCE [13], and Cardial pottery is found in parts of Iberia by ~5500 cal BCE [14]. Northwestern France was not reached until ~4500 BCE, and the British Isles and southern Scandinavia (Funnel Beakers) not until ~4000 BCE [13,15].
In addition to the two land routes, long-distance sea travel originating in northwest Italy has been proposed to account for the presence of early (before ~5600 cal BCE) Neolithic coastal sites in Iberia [16].
As noted above, early farming was introduced into Europe by migrants from Anatolia. Here we consider interactions, in particular admixture, with the autochthonous hunter-gatherers as revealed by ancient DNA evidence. The European hunter-gatherers in question are known as western hunter-gatherers (WHG). There was genetic heterogeneity among the WHG, possibly as a result of isolation in different refugia during the Last Glacial Maximum (LGM) [12,17,18]. Admixture is usually viewed as occurring by the integration of local hunter-gatherers into a farming community, i.e., by introgression [12,13,15,17,19,20]. Ancient DNA studies indicate that the Anatolian farmers spread through much of Europe with limited initial admixture from the indigenous hunter-gatherers, rarely exceeding 10% during the early Neolithic [12,14,17]. In effect, the spread of farming occurred by demic diffusion [1,5].
Nevertheless, the resident hunter-gatherers continued to coexist with the incoming farmers [17,21–23], and not only in Scandinavia [15,24]. That is, hunter-gatherer societies did not completely disappear because of competitive exclusion. For example, of the two contemporaneous samples excavated from two sites of Neolithic Körös in Hungary, KO1 (5,650–5,780 cal BCE) is genetically classified as hunter-gatherer, whereas KO2 (5,570–5,710 cal BCE) clusters with later Neolithic individuals [22]. As another example, the Blätterhöhle site in Germany has a Late Neolithic (3986–2918 cal BCE) occupation, which has yielded many individuals belonging to Mesolithic and early Neolithic ancient mitochondrial lineages [21]. Dietary stable isotope analysis on these samples reveals an association with corresponding lifestyles (i.e., subsistence), which suggests the prolonged coexistence of hunter-gatherers and farmers in central Europe.
Ancient DNA can also be used to estimate inbreeding effective population size (roughly the number of reproducing individuals). Runs of homozygosity (ROH) are segments of homologous chromosomes in an individual that are homozygous (i.e., with identical DNA sequences). Smaller populations have more and longer ROH than larger populations; other factors such as admixture and inbreeding also affect ROH. Ceballos et al. [25] find very high levels of ROH in Palaeolithic and Mesolithic samples, whereas Neolithic samples show much reduced levels. Using this method, Günther et al. [26] estimate the approximate size of the mating-network to which the Mesolithic hunter-gatherer Loschbour (Luxembourg) or StoraFörvar12 (Sweden) belonged to have been ~3000, whereas that of the LBK farmer Stuttgart (southern Germany, ~5000 BCE) was >20000.
Ancient DNA studies yield evidence of possible differences in physical appearance between European hunter-gatherers and Anatolian farmers, specifically in their skin and eye colors, which may be relevant to the later discussion. SLC45A2 (rs16891982) and SLC24A5 (rs1426654) are the two loci (single nucleotide polymorphisms or SNPs) with the strongest effects on skin color in present-day Europeans, where the ancestral and derived alleles are associated with dark and light skin color, respectively. Similarly, the derived allele of HERC2 (rs12913832) is the primary determinant of lighter eye color (e.g., blue).
An early study by Olalde et al. [27] found that Mesolithic La Braña 1 (northwest Spain, ~5000 BCE) was homozygous for the ancestral alleles of both SLC45A2 and SLC24A5, suggesting that this individual was dark-skinned. On the other hand, this individual was homozygous for the derived allele of HERC2, indicating that this individual was perhaps blue-eyed. Subsequent studies support a similar phenotypic combination for numerous other WHG [28–30]. Scandinavian hunter-gatherers (SHG) need to be distinguished from WHG, however, because six individuals from Motala (southern Sweden, 5900-5500 BCE) have predominantly derived alleles at SLC45A2 and SLC24A5 [31].
The derived allele of SLC24A5 was apparently fixed among 26 individuals of the Anatolian Neolithic (6400–6300 BCE; 21 of whom come from Barcin Höyük in northwest Turkey), whereas the derived allele of SLC45A2 occurred at low frequency [31]. Hence, these Anatolian farmers may have had an intermediate complexion. Feldman et al. [32] find that three of five Neolithic individuals from Boncuklu (possibly older than 8000 BCE, southern Turkey) were also homozygous for the derived allele of SLC24A5. Incidentally, Stuttgart is also homozygous for the derived allele of SLC24A5 [28], consistent with Anatolian ancestry. Feldman et al. [32] were also able to show that two individuals were homozygous and one was heterozygous for the derived allele of HERC2. Hence, Anatolian farmers may have been polymorphic for eye color.
Interestingly, an Epipaleolithic hunter-gatherer excavated from the neighboring site of Pınarbaşı (13642–13073 cal BCE) is homozygous for the ancestral allele of both SLC24A5 and HERC2 [32].
Let f(x,t), c(x,t), and h(x,t) denote the densities of the Anatolian farmers, converted farmers (European descent), and hunter-gatherers (also European descent), respectively, at position x and time t in an environmentally-homogeneous infinite one-dimensional space. Aoki [10] proposed the following reaction-diffusion model:
In Equations (3), the intrinsic rates of growth of the three populations, rf, rc, and rh, may differ, whereas the diffusion coefficient, D, is assumed to be the same for all three populations. The combined carrying capacity of Anatolian and converted farmers is K, and that of hunter-gatherers is L. Parameter e in the third term on the right-hand sides of Equations (3b) and (3c) represents the local conversion rate of hunter-gatherers to farmers, where consistency requires e · max(f + c) ≤ 1.
Equations (3a) extend the model formulated by Aoki et al. [9] by the addition of Lotka-Volterra competition terms between the farmers and the hunter-gatherers. Specifically, pf and pc represent the competitive effect of hunter-gatherers on the Anatolian farmers and converted farmers, respectively, and q denotes the competitive effect of farmers on hunter-gatherers (‘inter-subsistence’ competition). The ‘intra-subsistence’ competition coefficient between Anatolian and converted farmers has been set to 1 (i.e., f and c have equivalent effects in suppressing logistic growth in Equations (3a) and (3b). Equations (3) are identical to Equation 1 of Aoki [10], except for a minor notational change: pf replaces w and pc replaces p. All parameters are non-negative.
We pause to note three limitations of the model. First and most basic, Equations (3) assume an environmentally-homogeneous one-dimensional space with constant parameter values; this is limitation #1. Second, they assume asexual reproduction and a permanent distinction between the Anatolian and converted farmers. In reality, converted farmers may comprise individuals of varying degrees of admixture and/or of assimilation into farming society; this is limitation #2. Third, in contrast to the intrinsic growth rates and the carrying capacities for which (order-of-magnitude) estimates exist, the competition coefficients, pf, pc, q, and perhaps also the conversion (acculturation) rate, e, can only be assigned speculative values; this is limitation #3. It should be noted here that Joaquim Fort (and his colleagues) have done much theoretical modeling with an empirical basis, i.e., their models are formulated in terms of parameters that can be and are estimated from data (see Fort [33] for review).
One reviewer pointed out that estimates of the conversion rate are available. There are many alternative formulations of conversion, e.g., [34–36]. Conversion by horizontal transmission as modeled in Equations (3b) and (3c) is of the same basic form as in Sgaramella-Zonta and Cavalli-Sforza [7] and Rendine et al. [8]; their estimate of the conversion rate is of the order of 10-4 per generation. Fort [35] provides direct estimates based on short-term ethnographic observations of Christian missionaries in contact with a larger number of hunter-gatherers. His method yields an estimate of 1~10.9 hunter-gatherers converted by each farmer per generation, a figure that he notes would be applicable only at the leading edge of the wave-of-advance. In addition, it is not clear whether relations between missionaries and hunter-gatherers can be regarded as typical of what transpired in prehistoric encounters between farmers and hunter-gatherers.
In what follows, we proceed on the assumption that Equations (3) can yield a traveling wave solution for the spread of farming. Numerical work described later provides support for this claim.
We begin by noting the meanings of the equilibria without spatial structure (D = 0) listed in Appendix 1. E1 corresponds to total extinction; E2 (Equation A3) and E3 (Equation A4) to elimination of farmers and hunter-gatherers, respectively; E4 (Equation A5) to elimination of Anatolian farmers; and E5 (Equations A6) to coexistence of all three populations. Travelling wave solutions in space connect two—or in some cases more than two [37]—such equilibria, with a continuous transition in between (see Figure 1 for an example). For the initial conditions, it seems natural to set f =

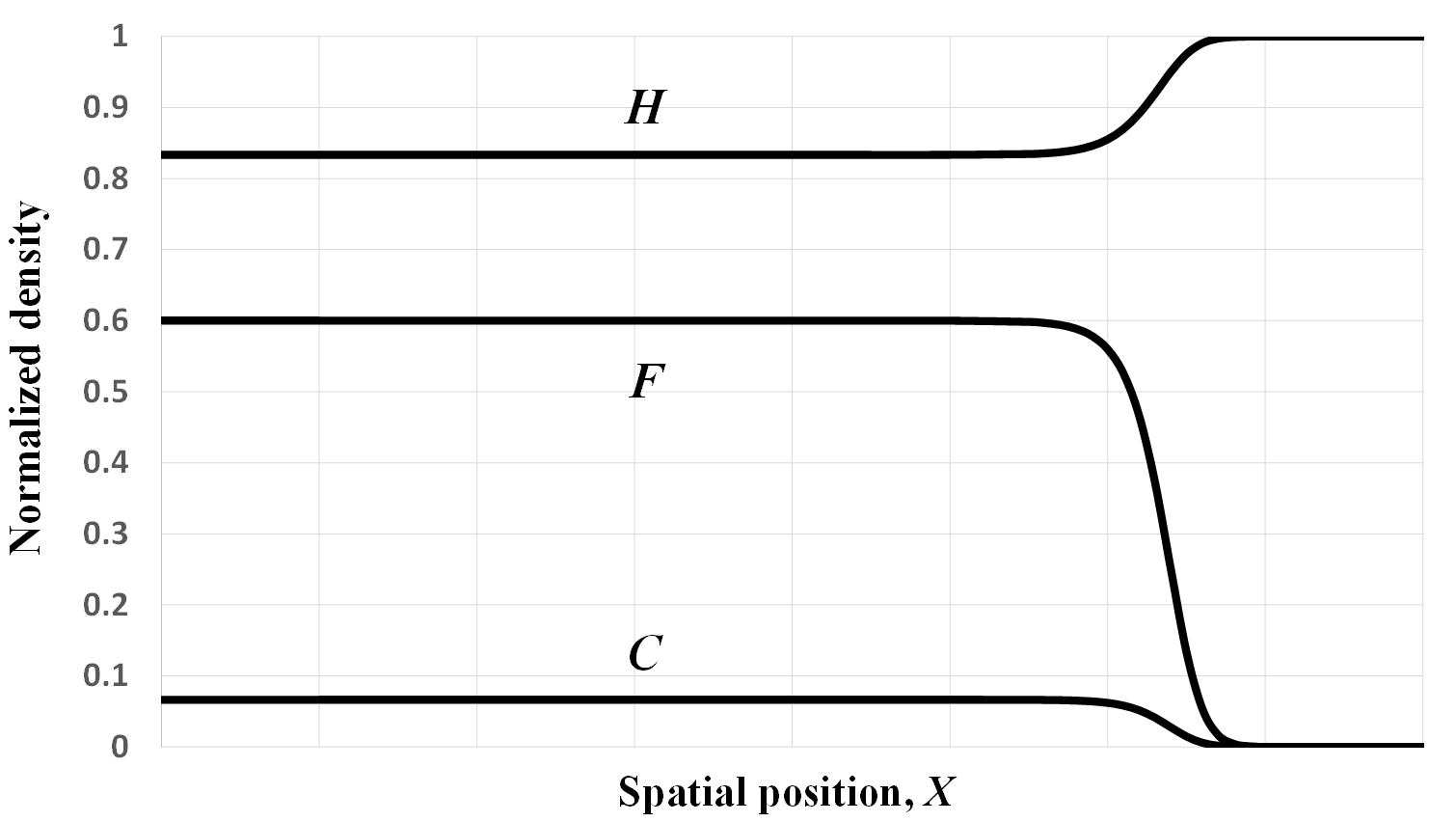
Figure 1. A numerically-simulated traveling wave solution. The vertical axis gives the normalized densities of the Anatolian farmers (labeled F), converted farmers (labeled C), and the autochthonous hunter-gatherers (labeled H). The wave-of-advance of farming comprises both Anatolian and converted farmers, the latter at relatively low density. Hunter-gatherers coexist with the farmers behind the wave front. See section 7 for details of the numerical method used. Parameter values are α = rf / rc = 1, β = rh / rc = 1, σ = eL/rc = 0.01, γ = eK/rh = 0.15, pf = 0.4, pc = 0.5, q = 0.1.
Equilibrium E2 should continue to hold in regions where the wave of advance has not yet penetrated. The empirical evidence reviewed in Section 3 suggests that the densities of the three populations behind the wave front of the spread of farming along either the Danubian or Mediterranean route should satisfy
Importantly, the conditions
are sufficient for the existence of E5 and necessary for its linear stability, where σ = eL/rc and γ = eK/rh [10]. Extensive numerical work suggests that inequalities Equations (4) are in fact necessary and sufficient for the existence and linear stability of equilibrium E5.
Inequalities Equations (4a) and (4c) together entail that the farmers will be present at below their carrying capacity, i.e.,
The (asymptotic) speed of this traveling wave is predicted to be either
or
whichever is greater (Appendix 2). Equations (5) are obtained from the non-dimensional Equations (A9) by multiplying by
Neither E2, E3, nor E4 can coexist with E5 as a linearly stable equilibrium. This suggests that, when E5 exists and is linearly stable, a monostable traveling wave will be formed in which the spatial region where E5 holds expands and the region where E2 holds contracts. It also entails that the region where E3 holds will eventually be eliminated, i.e., populations of European descent will invade Anatolia, which is not supported by data. This constitutes limitation #4 of the model.
Traveling wave solutions corresponding to cultural diffusion (a waveform connecting E4 to E2) are also possible. See Aoki [10] for details of the above including proofs.
Equations (3) are solved numerically in the following way. First, the independent variables, t and x, and the dependent variables, f, c, and h, are transformed to non-dimensional form (Appendix 2). This procedure yields Equations (A8), where each of the corresponding dependent variables, F, C, and H, equals 1 at carrying capacity. Equations (A8) have the merit that the number of independent parameters is reduced compared to Equations (3).
Second, time and space are discretized, and Equations (A8) are converted to difference equations, which are solved on a finite space lattice comprising 801 points (0 ≤ i ≤ 800). The non-dimensional time and space increments are set to ΔT = 0.005 and
Figure 1 illustrates one traveling wave solution obtained with this procedure, which constitutes a ‘snapshot’ of the spatial distribution of the three population densities at one moment in time. We assume here that the intrinsic growth rates are the same for all three populations, i.e., rf = rc = rh, which entails α = β = 1 in the non-dimensional coordinates. We further set the carrying capacity of farmers to 15 times that of hunter-gatherers [38], which is more conservative than the 50-fold difference assumed by Ammerman and Cavalli-Sforza [5]. The other parameter values have been chosen to satisfy the inequalities in Equations (4) and so that σ/(pc − pf) = 0.1 (see below).
As expected, we observe an advancing wave comprising both Anatolian and converted farmers. Moreover, hunter-gatherers coexist with the farmers behind the wave front. These features are consistent with the ancient DNA and archaeological evidence discussed above.
If we assume an intrinsic growth rate of ~4% per year (e.g., Ammerman and Cavalli-Sforza [4]; Zahid et al. [39] but see correction), then the traveling wave represents the situation ~2200 years (≈ 87/0.04, see below) after start of expansion. The rate of advance obtained from the numerical work is 1.518, which is in moderately good agreement with the non-dimensional speed 1.549 predicted by Equation (A9a). The speed in real time and space is
Toward the left of Figure 1, we can see an incursion into Anatolia of populations of European descent (i.e., C and H are both positive), which as previously noted is a limitation of the current model. Ironically, the simple Fisher model Equation (1) does not exhibit such an anomaly.
The average fraction of indigenous hunter-gatherer admixture in a farming society at location x and time t is given by c(x,t)/[f(x,t) + c(x,t)] provided the denominator is positive. This quantity can be regarded as the average of ancestry estimates for individuals obtained in genomic studies. We consider how this admixture fraction may vary in space at fixed times. When the traveling wave has been formed, Equations (A6a) and (A6b) entail that it equals σ/(pc − pf) at the equilibrium E5, in both the untransformed and transformed variables (Appendix 2). Let us define the average admixture fraction in terms of the transformed variables,
For the baseline example in the previous section, Figure 2a shows that A(X,87) = σ/(pC − pF) = 0.1 uniformly in space (0 ≤ X ≲ 179). Here, T ≈ 87 is the time in transformed variables prior to which the traveling wave is judged to have been formed, i.e., H ≅ 0.95 at i = 640. As argued above, this corresponds to ~2200 years given an intrinsic growth rate of ~4% per year. The prediction of spatial uniformity may not be inconsistent with the ancient DNA evidence, because Lipson et al. [17] (their Figure 2a) does not suggest significant spatial variation in hunter-gatherer ancestry proportions in the early Neolithic. Specifically, for the contemporaneous data circa 5000 BCE, there is considerable overlap in the hunter-gatherer ancestry proportions among samples from Hungary, the LBK, and Iberia. However higher proportions of hunter-gatherer ancestry have recently been reported from France [40] and Portugal [12], for example.
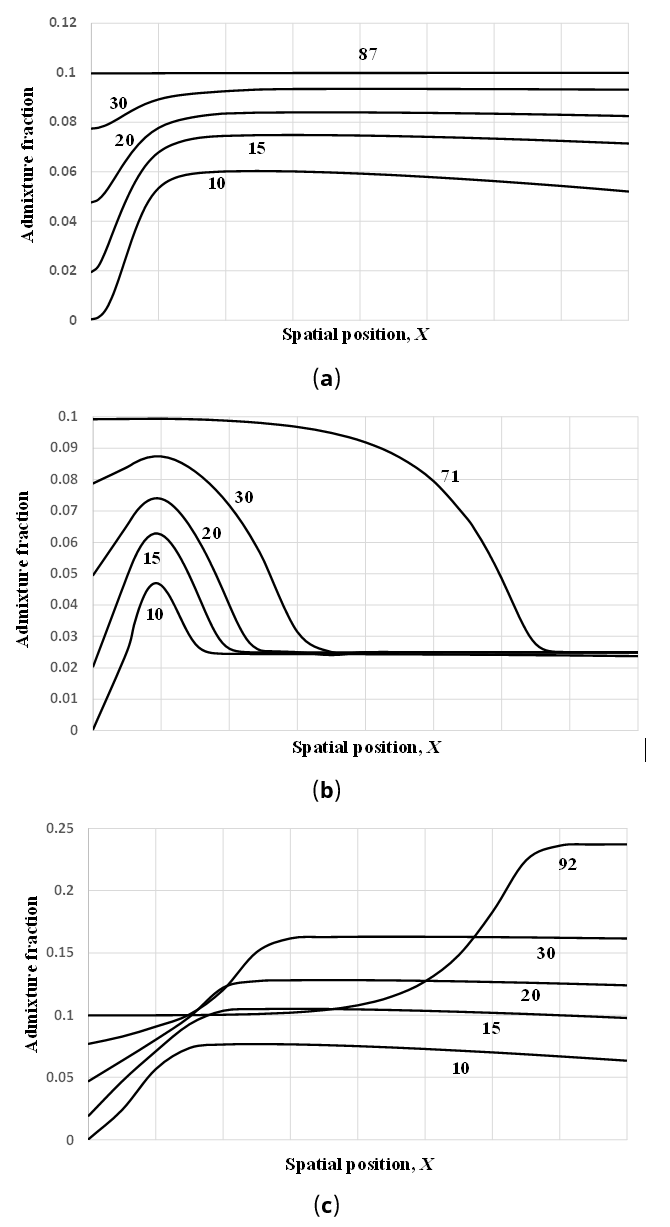
Figure 2. Average admixture/introgression fraction as a function of spatial position and time. The curves are labeled with the transformed time, T = rct, since beginning of expansion. (a) Parameter values are the same as in Figure 1. (b) Parameter values same except α = 1.5. (c) Parameter values same except α = 0.9.
On the other hand, Figure 2a may be considered incongruous in that it shows A(X,87) = 0.1 ahead of the wave front, where presumably farmers have not yet arrived. This is an artifact of the model attributable to bidirectional diffusion, which we list as possible limitation #5. However, small numbers of Anatolian and converted farmers may have migrated beyond the wave front, and the admixture fraction may be construed as reflecting a ratio of low densities.
Parameter α = rf/rc is the ratio of the intrinsic growth rates of Anatolian and converted farmers. Intuitively, we expect it to have a significant effect on A(X,T). If we set α = 1.5 in the baseline numerical example above without altering the other parameters, we find that the admixture fraction is relatively low at the wave front of the traveling wave solution (~0.03 when T ≈ 71) and approaches the equilibrium value of 0.1 in its wake (Figure 2b). If α = 0.9 < 1 in contrast, the admixture fraction is relatively high at the wave front (~0.22 when T ≈ 92) (Figure 2c). At the wave front where the densities of Anatolian and converted farmers are both low, the population with the larger intrinsic growth rate is expected to increase more rapidly, and hence these results are reasonable.
Figures 2a, 2b, 2c also show how A(X,T) varies in space in the initial phases of the spread of farming before the traveling wave has been established, i.e., at the transformed times T ≈ 10, 15, 20, 30. Spatial variation in the average admixture fraction is especially pronounced in Figure 2b. Figure 2c illustrates a case where A(X,T) does not increase monotonically in time at fixed points in space. The latter figure also shows the eventual formation at transformed time T ≈ 92 (~2300 years if rc = 4% per year) of a geographical gradient in the admixture fraction (higher in the west), in qualitative agreement with Rivollat et al. [40] and Allentoft et al. [12].
The three-population model originally proposed by Aoki et al. [9] did not include inter-subsistence competition, i.e., it is a special case of Equations (3) with pf = pc = q = 0. The only spatially-homogeneous equilibrium at which farmers and hunter-gatherers can coexist in this reduced model is:
This equilibrium exists and is linearly stable if and only if γ < 1 [9]. Note that Anatolian farmers are absent at this equilibrium (
Consider a traveling wave solution connecting this equilibrium with equilibrium E2 [hunter-gatherers only, Equation (A3)], where the latter is always unstable given pf = pc = q = 0. When γ < 1, one therefore expects a traveling wave solution in which the Anatolian farmers are eliminated and farming spreads by cultural diffusion. For extreme parameter sets, however, the Anatolian farmers may continue to be present at high density in a localized region at the wave front. Appendix Figure A1 illustrates such an outcome at T ≈ 55 (~1376 years if rc = 4% per year). In this example, α = 1.5, β = 1.0, and σ = γ = 0.1, which entails that the carrying capacities of farmers and hunter-gatherers are equal. The heuristically-predicted speed in real time and space is
It is debatable whether this peaked distribution of Anatolian farmers can be said to represent demic diffusion. In addition, the peaked waveform may be a transient phenomenon. This possibility was tested (result not shown) by employing a longer space lattice of 1601 points (0 ≤ i ≤ 1600). At non-dimensional time T ≈ 114 (~2851 years if rc = 4% per year), a pronounced peak of the same height and same width as in Appendix Figure A1 is still found to exist. Hence, Appendix Figure A1 may represent a bona fide traveling wave solution.
Patterson et al. [41] have introduced environmental heterogeneity (recall limitation #1) and other generalizations into the model of Aoki et al. [9]. They find that Near Eastern and converted farmers can coexist in regions favorable to farmers, but note that “no significant population of initial (Near Eastern) farmers survives there”.
Archaeologists have investigated changes in the relative sizes of populations over time by the method of the summed probability distribution (SPD) [42,43]. This method is based on the assumption that the number of reported radiocarbon dates from all known sites in a given area and given time window can serve as an approximate proxy for the corresponding relative population size. SPD is a plot of these numbers over the period of interest.
Applying this method to the Neolithic transition in various regions of Europe, Shennan et al. [44] identified a pattern of significant population growth with the appearance of farming followed by population collapse. They tentatively attributed this observation to endogenous causes such as rapid population growth to unsustainable levels. Silva and Vander Linden [45], on the other hand, interpret this boom-and-bust pattern as being caused by migration, with a boom reflecting the arrival and a bust the departure of a large group of people.
The form of a traveling wave may be monotone or peaked, e.g., Figure 1 shows that F, C, and H are each monotone decreasing or increasing in space. Aoki et al. [9] argued that a traveling wave of Near Eastern and converted farmers combined may be peaked under certain conditions. However, the peaks as a function of time predicted by numerical work are not nearly as prominent as the SPD studies suggest. Hence, the current model cannot explain the boom-and-bust pattern, which is limitation #6.
A consequence of Equations (3) is that the Anatolian farmers, converted farmers, and hunter-gatherers can coexist stably behind the Neolithic wave front—which is what the ancient DNA studies clearly imply—only if pc > pf + σ (Equation 4b). This means that the resident hunter-gatherers must behave more competitively toward the converted than the Anatolian farmers. From a theoretical standpoint, this prediction is understandable, because an increase in the density of converted farmers by acculturation should be balanced by their decrease by competitive exclusion. However, the empirical question remains as to why the hunter-gatherers would have discriminated between the Anatolian and converted farmers in this way.
Generally speaking, there are two contrasting views on how farmers (Near Eastern or converted) interacted with the resident hunter-gatherers that they encountered in Europe. Archaeological evidence for the LBK suggests either mutual avoidance in land use [46] or violent conflict [47]. Hence, excess competition directed toward converted farmers may mean less avoidance or more conflict.
The first generation converted farmers were perhaps women who married into the farming communities [48], in which case conflict seems unlikely. Nevertheless, transition to a farming lifestyle may have been gradual, and second and later generation converted farmer males may have hunted in their ancestral lands, or at the “forest-field edge” where wild animals may gather [49], resulting in resource competition. Another highly speculative possibility is that most (dark-skinned, blue-eyed) hunter-gatherers may have kept their distance from the phenotypically divergent (comparatively light-skinned, brown-eyed) Anatolian farmers.
Lucquin et al. [50] compared the diets of early farmers and Mesolithic hunter-gatherer-fishers in northern Europe, as revealed by organic residues in pottery. The hunter-gatherer-fishers received dairy products from the farmers, and the farmers utilized aquatic resources, which is interpreted as cooperative interaction. However, it is also noted that, in contrast to hunter-gatherers who may have been unable to independently transition to farming, farmers were easily able to adopt foraging strategies. If the converted farmers contributed to this asymmetry, they may have provoked competition from the hunter-gatherers.
Not applicable.
Not applicable.
Not applicable.
None.
The author has declared no competing interests exist.
I am indebted to Naoyuki Takahata, Yukiko Kawanishi, and one reviewer for directing me to relevant literature. I also thank the editors and both reviewers for valuable comments on the paper.
f, c, and h are now functions of time t only, which satisfy the ordinary differential equations
The subscript t here indicates ordinary differentiation with respect to time.
The equilibria of Equation (A1) are
and
where σ = eL/rc and γ = eK/rh.
In Appendix 2 of Aoki [10], it is shown that conditions
are sufficient for the existence of equilibrium E5 and necessary for its linear stability (all eigenvalues are negative or have negative real part). Moreover, neither E2, E3, nor E4 can coexist with E5 as a linearly stable equilibrium.
Following standard practice [51], we transform Equations (3) to non-dimensional form
Here, T = rct,
Heuristic phase space analysis [6] can be used to predict the (minimum) speed of the wave-of-advance. Set Z = X − vT, where v is this speed, and assume that the densities are functions of Z, rather than of X and T separately. By conducting local stability analysis in the six-dimensional phase space of F,FZ, C, CZ, H and HZ, it is possible to obtain necessary conditions for non-negative wave-of-advance solutions to exist, which in turn yield predictions of the speed. Applying this procedure to Equations (A8) gives [10]
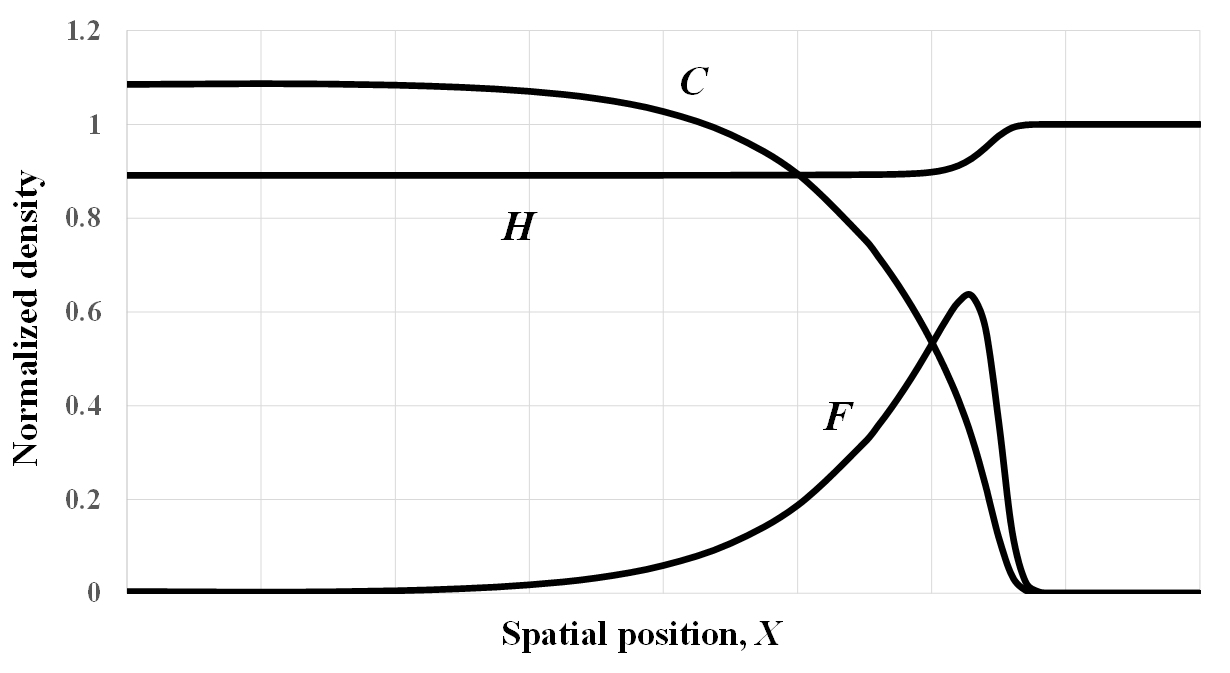
Figure A1. A numerically-simulated traveling wave solution with parameter values α = rf / rc = 1.5, β = rh / rc = 1, σ = eL/rc = 0.1, γ = eK/rh = 0.1, pf = 0, pc = 0, q = 0. Note that the three interspecific competition parameters have been set to 0.
| 1. | Ammerman AJ, Cavalli-Sforza LL. Measuring the rate of spread of early farming in Europe. Man. 1971;6:674-688. [Google Scholar] [CrossRef] |
| 2. | Ammerman AJ. The transition to early farming in Europe. In: Pardo-Gordó S, Bergin S, editors. Simulating transitions to agriculture in prehistory. Switzerland: Springer Nature; 2021; pp. 225–253. |
| 3. | Kendall DG. A form of wave propagation associated with the equation of heat conduction. Proc Camb Phil Soc. 1948;44:591-594. [Google Scholar] [CrossRef] |
| 4. | Skellam J. Random dispersal in theoretical populations. Biometrika. 1951;38:196-218. [Google Scholar] [CrossRef] |
| 5. | Ammerman AJ, Cavalli-Sforza LL. A population model for the spread of early farming in Europe. In: Renfrew C, editor. The explanation of culture change. London: Duckworth; 1973; pp. 343–357. |
| 6. | Fisher RA. The wave of advance of advantageous genes. Ann Eugen. 1937;7:355-369. [Google Scholar] |
| 7. | Sgaramella-Zonta L, Cavalli-Sforza LL. A method for the detection of a demic cline. In: Morton NE, editor. Genetic structure of human populations. Honolulu: Univ Hawaii Press; 1973; pp. 128–135. |
| 8. | Rendine S, Piazza A, Cavalli-Sforza LL. Simulation and separation by principal components of multiple demic expansions in Europe. Am Nat. 1986;128:681-706. [Google Scholar] [CrossRef] |
| 9. | Aoki K, Shida M, Shigesada N. Travelling wave solutions for the spread of farmers into a region occupied by hunter-gatherers. Theor Popul Biol. 1996;50:1-17. [Google Scholar] [CrossRef] |
| 10. | Aoki K. A three-population wave-of-advance model for the European early Neolithic. PLoS One. 2020;15(5):e0233184. [Google Scholar] [CrossRef] |
| 11. | Pinhasi R, Fort J, Ammerman AJ. Tracing the origin and spread of agriculture in Europe. PLoS Biol. 2005;3(12):e410. [Google Scholar] [CrossRef] |
| 12. | Allentoft ME, Sikora M, Refoyo-Martínez A, Irving-Pease EK, Fischer A, Barrie W, et al. Population genomics of post-glacial western Eurasia. Nature. 2024;625:301-311. [Google Scholar] [CrossRef] |
| 13. | Furholt M. Mobility and social change: understanding the European Neolithic period after the archaeogenetic revolution. J Archaeol Res. 2021;29:481-535. [Google Scholar] [CrossRef] |
| 14. | Valdiosera C, Günther T, Vera-Rodríguez JC, Ureña I, Iriarte E, Rodríguez-Varela R, et al. Four millennia of Iberian biomolecular prehistory illustrate the impact of prehistoric migrations at the far end of Eurasia. Proc Natl Acad Sci USA. 2018;115:3428-3433. [Google Scholar] [CrossRef] |
| 15. | Skoglund P, Malmström H, Omrak A, Raghavan M, Valdiosera C, Günther T, et al. Genomic diversity and admixture differs for stone-age Scandinavian foragers and farmers. Science. 2014;344:747-750. [Google Scholar] [CrossRef] |
| 16. | Isern N, Zilhão J, Fort J, Ammerman AJ. Modeling the role of voyaging in the coastal spread of the Early Neolithic in the West Mediterranean. Proc Natl Acad Sci USA. 2017;114:897-902. [Google Scholar] [CrossRef] |
| 17. | Lipson M, Szécsényi-Nagy A, Mallick S, Pósa A, Stégmár B, Keerl V, et al. 2017. Parallel paleogenomic transects reveal complex genetic history of early European farmers. Nature. 2017;551:368-372. [Google Scholar] [CrossRef] |
| 18. | Villalba-Mouco V, van de Loosdrecht MS, Posth C, Mora R, Martínez-Moreno J, Rojo-Guerra M, et al. Survival of late Pleistocene hunter-gatherer ancestry in the Iberian Peninsula. Curr Biol. 2019;29:1169-1177. [Google Scholar] [CrossRef] |
| 19. |
Nikitin AG, Stadler P, Kotova N, Teschler-Nicola M, Price DT, Hoover J, et al. Interactions between earliest |
| 20. | Wang K, Prüfer K, Krause-Kyora B, Childebayeva A, Schuenemann VJ, Coia V, et al. High-coverage genome of the Tyrolean Iceman reveals unusually high Anatolian farmer ancestry. Cell Genomics. 2023;3:100377. [Google Scholar] [CrossRef] |
| 21. | Bollongino R, Nehlich O, Richards MP, Orschiedt J, Thomas MG, Sell C, et al. 2000 years of parallel societies in stone age central Europe. Science. 2013;342:479-481. [Google Scholar] [CrossRef] |
| 22. | Gamba C, Jones ER, Teasdale MD, McLaughlin RL, Gonzalez-Fortes G, Mattiangeli V, et al. Genome flux and stasis in a five millennium transect of European prehistory. Nat Commun. 2014;5:5257. [Google Scholar] [CrossRef] |
| 23. | Mathieson I, Alpaslan-Roodenberg S, Posth C, Szécsényi-Nagy A, Rohland N, Mallick S, et al. The genomic history of southeastern Europe. Nature. 2018;555:197-203. [Google Scholar] [CrossRef] |
| 24. | Malmström H, Gilbert MTP, Thomas MG, Brandström M, Storå J, Molnar P, Andersen PK, et al. Ancient DNA reveals lack of continuity between Neolithic hunter-gatherers and contemporary Scandinavians. Curr Biol. 2009;19:1758-1762. [Google Scholar] [CrossRef] |
| 25. | Ceballos FC, Joshi PK, Clark DW, Ramsay M, Wilson JF, et al. Runs of homozygosity: windows into population history and trait architecture. Nat Rev Genet. 2018;19:220-234. [Google Scholar] [CrossRef] |
| 26. | Günther T, Malmström H, Svensson EM, Omrak A, Sánchez-Quinto F, Kılınç GM, et al. Population genomics of Mesolithic Scandinavia: investigating early postglacial migration routes and high-latitude adaptation. PLoS Biol. 2018;16(1):e2003703. [Google Scholar] [CrossRef] |
| 27. | Olalde I, Allentoft M, Sánchez-Quinto F, Santpere G, Chiang CWK, DeGiorgio M, et al. Derived immune and ancestral pigmentation alleles in a 7,000-year-old Mesolithic European. Nature. 2014;507:225-228. [Google Scholar] [CrossRef] |
| 28. | Lazaridis I, Patterson N, Mittnik A, Renaud G, Mallick S, Kirsanow K, et al. Ancient human genomes suggest three ancestral populations for present-day Europeans. Nature. 2014;513:409-413. [Google Scholar] [CrossRef] |
| 29. | Fu Q, Posth C, Hajdinjak M, Petr M, Mallick S, Fernandes D, et al. The genetic history of Ice Age Europe. Nature. 2016;534:200-205. [Google Scholar] [CrossRef] |
| 30. | Posth C, Yu H, Ghalichi A, Rougier H, Crevecoeur I, Huang Y, et al. Palaeogenomics of Upper Palaeolithic to Neolithic European hunter-gatherers. Nature. 2023;615:117-126. [Google Scholar] [CrossRef] |
| 31. | Mathieson I, Lazaridis I, Rohland N, Mallick S, Patterson N, Roodenberg SA, et al. Genome-wide patterns of selection in 230 ancient Eurasians. Nature. 2015;528:499-503. [Google Scholar] [CrossRef] |
| 32. | Feldman M, Fernández-Domínguez E, Reynolds L, Baird D, Pearson J, Hershkovitz I, et al. Late Pleistocene human genome suggests a local origin for the first farmers of central Anatolia. Nat Commun. 2019;10:1218. [Google Scholar] [CrossRef] |
| 33. | Fort J. Prehistoric spread rates and genetic clines. Hum Popul Genet Genomics. 2022;2(2):0003. [Google Scholar] [CrossRef] |
| 34. | Barbujani G, Sokal RR, Oden NL, et al. Indo-European origins: a computer simulation test of five hypotheses. Am J Phys Anthropol. 1995;96:109-132. [Google Scholar] [CrossRef] |
| 35. | Fort J. Synthesis between demic and cultural diffusion in the Neolithic transition in Europe. Proc Natl Acad Sci USA. 2012;109:18669-18673. [Google Scholar] [CrossRef] |
| 36. | Isern N, Fort J, de Rioja VL, et al. The ancient cline of haplogroup K implies that the Neolithic transition in Europe was mainly demic. Sci Rep. 2017;7:11229. [Google Scholar] [CrossRef] |
| 37. | Wakano JY, Gilpin W, Kadowaki S, Feldman MW, Aoki K. Ecocultural range-expansion scenarios for the replacement or assimilation of Neanderthals by modern humans. Theor Popul Biol. 2018;119:3-14. [Google Scholar] [CrossRef] |
| 38. | Harper KN, Armelagos GJ. Genomics, the origins of agriculture, and our changing microbe-scape: time to revisit some old tales and tell some new ones. Am J Phys Anthropol. 2013;57:135-152. [Google Scholar] [CrossRef] |
| 39. | Zahid HJ, Robinson E, Kelly RL. Agriculture, population growth, and statistical analysis of the radiocarbon record. Proc Natl Acad Sci USA. 2015;113:931-935. [Google Scholar] [CrossRef] |
| 40. | Rivollat M, Jeong C, Schiffels S, Küçükkalıpçı İ, Pemonge M, Rohrlach AB, et al. Ancient genome-wide DNA from France highlights the complexity of interactions between Mesolithic hunter-gatherers and Neolithic farmers. Sci Adv. 2020;6:eaaz5344. [Google Scholar] [CrossRef] |
| 41. | Patterson MA, Sarson GR, Sarson HC, Shukurov A. Modelling the Neolithic transition in a heterogeneous environment. J Archaeol Sci. 2010;37:2929-2937. [Google Scholar] [CrossRef] |
| 42. | Rick JW. Dates as data: an examination of the Peruvian Preceramic radiocarbon record. Am Antiq. 1987;52:55-73. [Google Scholar] [CrossRef] |
| 43. | Williams A. The use of summed probability distributions in archaeology: a review of methods. J Archaeol Sci. 2012;39:578-589. [Google Scholar] [CrossRef] |
| 44. | Shennan S, Downey S., Timpson A, Edinborough K, Colledge S, Kerig T, et al. Regional population collapse followed initial agriculture booms in mid-Holocene Europe. Nat Commun. 2013;4:2486. [Google Scholar] [CrossRef] |
| 45. | Silva F, Vander Linden M. Amplitude of travelling front as inferred from 14C predicts levels of genetic admixture among European early farmers. Sci Rep. 2017;7:11985. [Google Scholar] [CrossRef] |
| 46. | Vanmontfort B. Forager-farmer connections in an ‘unoccupied’ land: first contact on the western front of LBK territory. J Anthropol Archaeol. 2008;27:149-160. [Google Scholar] [CrossRef] |
| 47. |
Golitko M, Keeley LH. Beating ploughshares back into swords: warfare in the |
| 48. | Bentley RA, Layton RH, Tehrani J, et al. Kinship, marriage, and the genetics of past human dispersals. Hum Biol. 2009;81:159-179. [Google Scholar] [CrossRef] |
| 49. | Peterson JT. The effect of farming expansion on hunting. Philippine Sociol Rev. 1982;30:33-50. [Google Scholar] |
| 50. | Lucquin A, Robsonet HK, Oras E, Lundy J, Moretti G, Carretero LG, et al. The impact of farming on prehistoric culinary practices throughout northern Europe. Proc Natl Acad Sci USA. 2023;120(43):e2310138120. [Google Scholar] [CrossRef] |
| 51. | Murray JD. Mathematical biology. Berlin: Springer; 1989. [Google Scholar] |

![]()
Copyright © 2026 Pivot Science Publications Corp. - unless otherwise stated | Terms and Conditions | Privacy Policy




