
Human Population Genetics and Genomics ISSN 2770-5005
Human Population Genetics and Genomics 2026;6(1):0003 | https://doi.org/10.47248/hpgg2606010003
Perspective Open Access
New insights from old DNA into the colonization of the Pacific: Two case studies
Mark Stoneking


Academic Editor(s): Joshua Akey, Carina Schlebusch
Received: Oct 31, 2025 | Accepted: Feb 13, 2026 | Published: Feb 26, 2026
© 2026 by the author(s). This is an Open Access article distributed under the Creative Commons License Attribution 4.0 International (CC BY 4.0) license, which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is correctly credited.
Cite this article: Stoneking M. New insights from old DNA into the colonization of the Pacific: Two case studies. Hum Popul Genet Genom. 2026;6(1):0003. https://doi.org/10.47248/hpgg2606010003
This paper is based on a keynote lecture given at the Wellcome Trust meeting on Human Evolution: From Fossils to Ancient Genomes in April 2025. After giving some background on the colonization of the Pacific, I describe the results of two of our studies of ancient DNA. The first used ancient DNA from Eastern Indonesia to address questions about the timing of the admixture between indigenous Papuan groups and incoming Austronesian groups, and showed that this admixture likely occurred soon after the arrival of Austronesians in the region. However, there were unexpected differences in the ancestry of ancient samples from northern Eastern Indonesia (i.e., the North Moluccas) vs. southern Eastern Indonesia (i.e., the Nusa Tenggaras): the latter had ancestry from mainland Southeast Asia that was absent in the former. The second study investigated the origin and relationships of the early colonists of Guam, in the Marianas Archipelago, and found that the most probable origin was from the Philippines, in agreement with linguistic evidence and some interpretations of the archaeological (pottery) evidence, but contrary to the results of computer simulations of voyaging. These studies illustrate the power of ancient DNA analyses to address questions about population history, as well as to provide novel insights that were not realized from studies of modern populations.
Keywordsancient DNA, Pacific colonization, Austronesian–Papuan admixture, Guam early settlers
This work is a written version of a keynote lecture given, with the same title, at the Wellcome Trust meeting on Human Evolution: From Fossils to Ancient Genomes, held at the Wellcome Genome Campus from 28-30 April 2025. There are three parts to this paper: first, some background information concerning the colonization of the Pacific; second, a discussion of a study of ancient DNA from Eastern Indonesia [1]; and third, a discussion of a study of ancient DNA from Guam [2]. I stress that this is not a comprehensive review of the literature pertaining to ancient DNA and the colonization of the Pacific; the reader interested in this topic should consult recent reviews [3,4]. Or, for the non-expert in genomics, I can recommend the relevant chapter in the recently-published second edition of my book, An Introduction to Molecular Anthropology [5].
The Pacific is a vast region encompassing roughly half the globe. The Polynesian Triangle alone (that is, the triangle with New Zealand, Rapa Nui, and Hawaii at its vertices) encompasses over 16 million km2, more than twice the size of the continental United States (Figure 1). Traditionally, Oceania (Figure 1) is divided into three main cultural areas, namely Melanesia (the dark islands), Polynesia (the many islands), and Micronesia (the tiny islands). However, these cultural areas do not reflect the colonization history of the region; for this purpose, a more informative distinction is made between Near Oceania (which consists of the main islands of Australia and New Guinea, extending through the Bismarck Archipelago and the main chain of the Solomon Islands) and Remote Oceania (all of the other islands east and north of Near Oceania, see Figure 1). Humans reached Near Oceania by at least 40-45 thousand years ago (ky) [3,6], when sea levels were much lower and Australia and New Guinea were connected as a single landmass known as Sahul. The water crossings necessary to reach Sahul from Sunda (the continental Asian landmass that included the present islands of Sumatra, Java, and Borneo) at this time were all intervisible (meaning that voyagers would see some indication of land ahead of them before losing sight of land behind them) and hence did not require sophisticated boating technology or navigation skills. By contrast, to reach the various islands of Remote Oceania meant crossing several hundred to a few thousand kilometers of open ocean, and required more sophisticated boating technology (e.g., outrigger canoes) and navigational skills. Even so, Remote Oceania was colonized beginning about 3-3.5 kya, with the remotest islands reached by about 1 kya [7,8] – and this at a time when European ships did not dare venture very far out of sight of land.

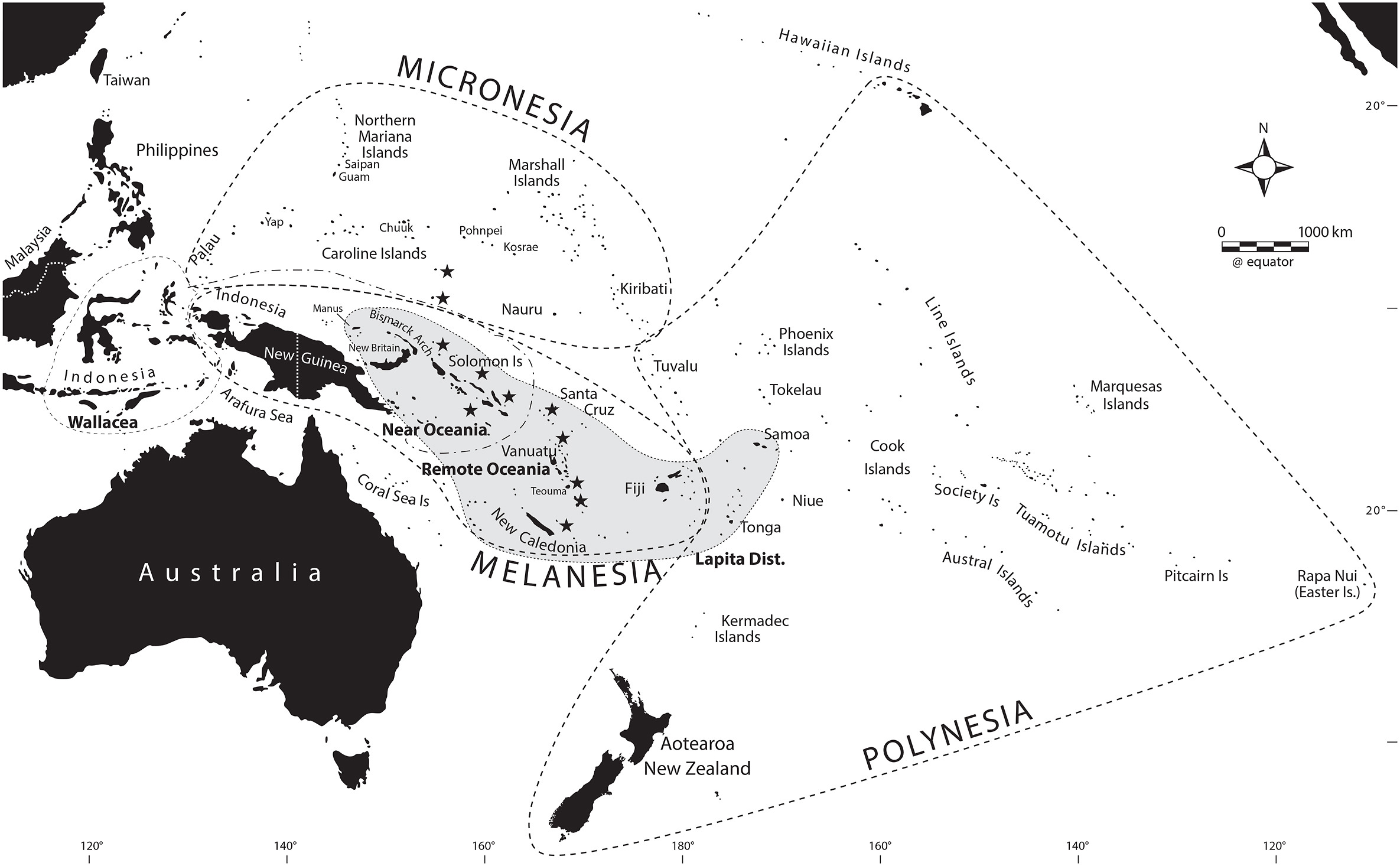
Figure 1. Map of Oceania, indicating the cultural areas of Melanesia, Micronesia, and Polynesia, and the boundary between Near and Remote Oceania. Reprinted with permission from [3].
Archaeological, linguistic, and genetic evidence all support two primary waves of colonization of Oceania, while leaving open the possibility of additional contact. The first is an early colonization of Near Oceania by at least 40-45 kya, and perhaps as much as 50-55 kya [6,9]. It is likely that this represents one of the earliest dispersals of modern humans; that is, after modern humans left Africa and mixed with Neandertals, the first migration from this non-African source population went via a southern route to Sahul [10]. The mountainous interior of New Guinea led to isolation of the groups living there, and promoted cultural, linguistic, and genetic diversity [3]; further details are provided in the next section. The second wave involved a migration of Austronesian-speaking rice and millet agriculturalists who left Taiwan (or possibly southern China) around 6 kya and island-hopped their way via the Philippines and Indonesia to the Bismarck Archipelago, spreading from there to Remote Oceania beginning around 3-3.5 kya [11,12]. The archaeological signature of the Austronesians is the characteristic dentate (stamped) pottery known as Lapita [12], while the languages are discussed in more detail in the next section. But within this broad framework there are many unresolved questions, and two of these are the subject of the two studies discussed here.
Eastern Indonesia, which comprises the islands east of Wallace's line and west of New Guinea, has long been recognized as a crossroads for human migrations between Southeast Asia and Near Oceania. This is evident in the languages, as both Papuan and Austronesian languages occur in the region. The Austronesian language family is one of the largest in the world, with some 1200 languages and over 350 million speakers; all of the languages of Taiwan and the Philippines, most of the languages of Indonesia, many of the languages of coastal and island Melanesia, and all of the languages of Remote Oceania are Austronesian languages [13] – Austronesians were even the first to get to Madagascar [14]. Both traditional and computational (phylogenetic) linguistic analyses strongly support a Taiwanese origin of Austronesian languages [13,15]. While Austronesian languages constitute a well-defined family (i.e., all Austronesian languages share particular features and can be reconstructed to a common ancestor called proto-Austronesian), by contrast so-called "Papuan" languages are mostly defined by what they are not: namely, they are indigenous languages spoken on New Guinea and a few nearby islands that are not Austronesian languages. The current situation is that Papuan languages comprise 23 distinct language families and several isolates [16], so even though I will refer to Papuan languages as a single entity, this underlying heterogeneity should be kept in mind (and in all likelihood similar heterogeneity underlies "Papuan" ancestry, even though I will also refer to Papuan ancestry as if it were a single ancestry). The general consensus is that the Papuan languages of Eastern Indonesia are a remnant of the languages that were spoken by the indigenous inhabitants of Eastern Indonesia prior to the arrival of Austronesians in the region, around 4 kya [15,17].
This view is supported by genetic studies, which have found a gradient of increasing Papuan-related ancestry and decreasing Austronesian-related ancestry as one moves from west to east through the region [18-20]. Presumably, the indigenous groups in the region (prior to the arrival of the Austronesians) were Papuan, and the gradient reflects the direction of spread of incoming Austronesian ancestry among the indigenous Papuan groups in the region. However, there is controversy over the timing of the genetic admixture between Papuan-related and Austronesian-related ancestry (Figure 2). Different methods for estimating the admixture date give different results, even when applied to the same datasets; some estimates are around 3-5 kya, whereas others are more recent, around 1-2 kya (Figure 2). Whether the actual date is around 3-5 kya vs. around 1-2 kya is not merely of academic interest, but rather has important implications for the timing of the interactions between the indigenous Papuan groups and the incoming Austronesian groups. If the true dates are around 3-5 kya, then this would suggest that the admixture occurred relatively soon after the arrival of the Austronesians, but if the true dates are more like 1-2 kya, then this would suggest that there was a long period of co-existence of the Austronesians and the indigenous Papuan groups (up to 1-2 ky) before they began admixing (this has been called the "getting to know you" hypothesis).
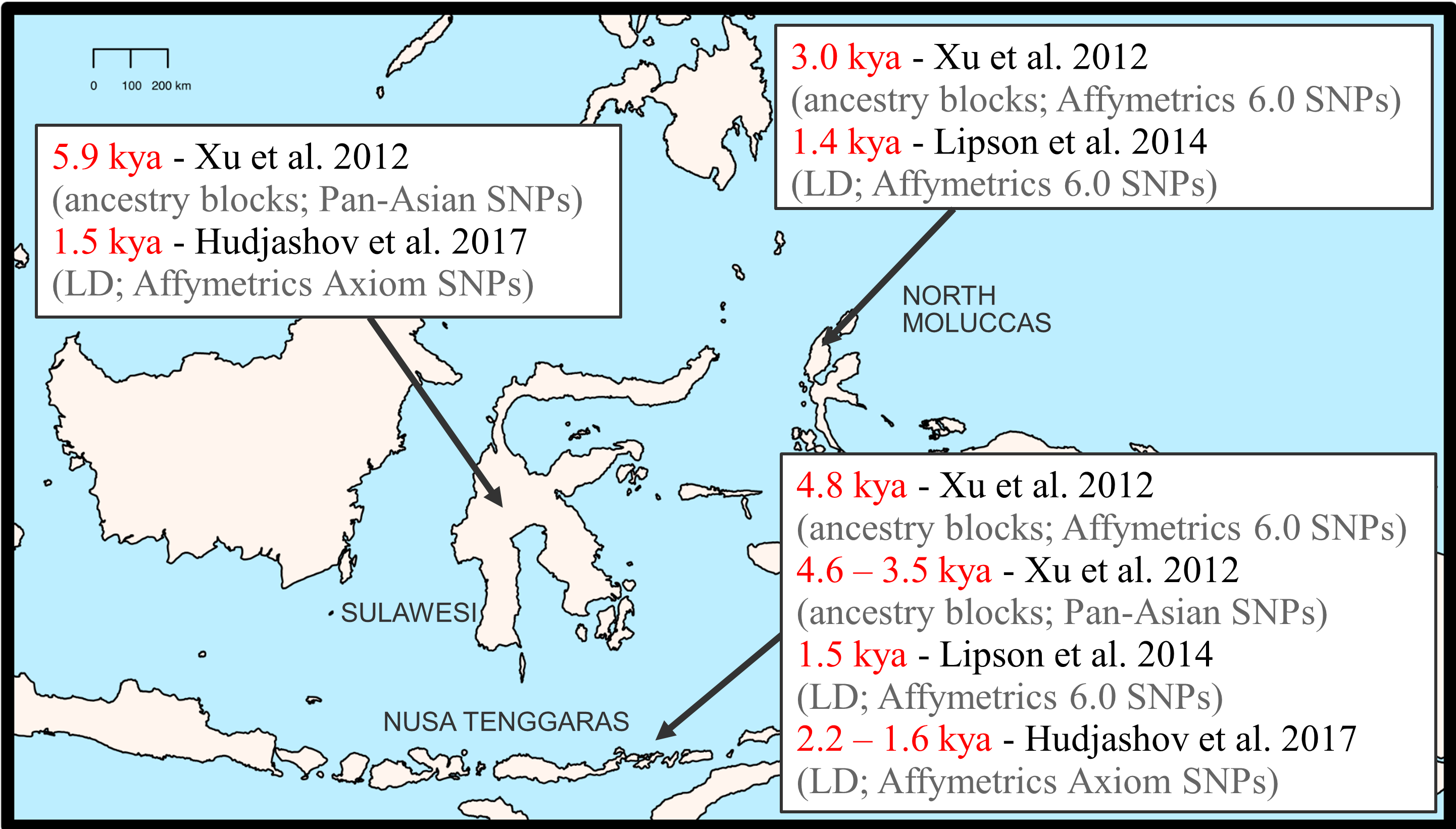
Figure 2. Widely-varying estimates of the admixture time between Austronesian-related and Papuan-related ancestry in Eastern Indonesia, from different studies that used different approaches (based on the size/number of ancestry blocks vs. LD) and different datasets (genotyping platforms) to estimate the admixture time.
What might help resolve this issue – or at least shed more light – would be ancient DNA data, ideally from a timespan that included the arrival of Austronesians in the region. In a collaboration with Johannes Krause and members of his group and with the assistance of several archaeologists working in the region [1], we were able to obtain genomic data from 16 individuals from several different sites across Eastern Indonesia (Figure 3A). They ranged in age from 0.25 – 2.6 kya, so alas none of the samples pre-dated the arrival of the Austronesians, but still they were old enough to tell us if admixture only began 1-2 kya vs. more than 3 kya.
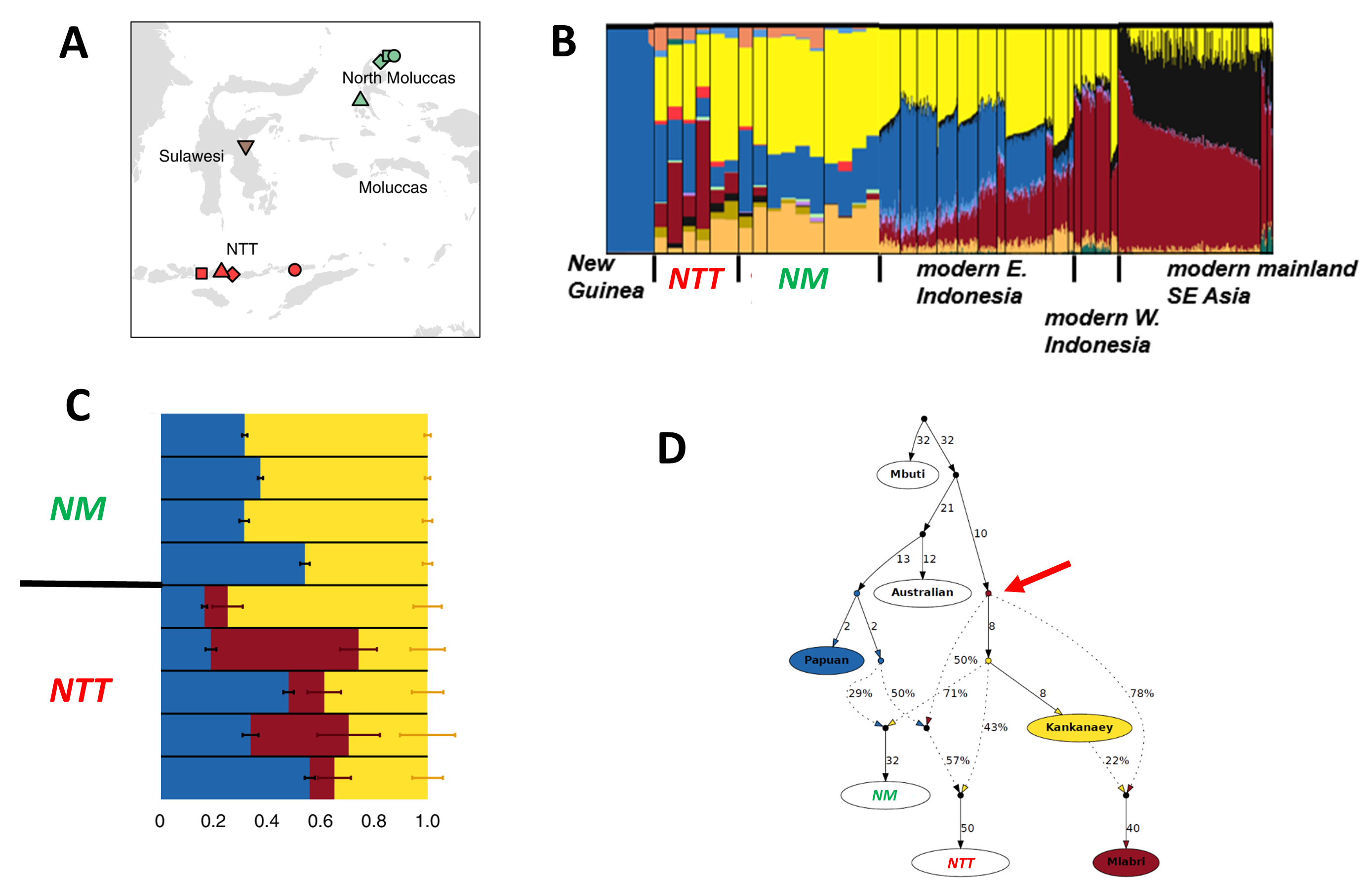
Figure 3. Analyses of ancient DNA from Eastern Indonesia. A: map of sampling locations. B: results of an ADMIXTURE analysis, focusing on the populations of interest. New Guinea exhibits a blue ancestry component that is also present in the ancient and modern samples from Eastern Indonesia, but not in modern samples from Western Indonesia or Mainland Southeast Asia. The yellow and peach ancestry components are of Austronesian origin. Notably, the red-brown ancestry component, which is present at highest frequency in modern samples from Mainland Southeast Asia, is also present in the ancient samples from the Nusa Tenggaras (NTT) but not in the ancient samples from the North Moluccas (NM). See [1] for further details. C: results of a qpAdm analysis. The NM samples can be successfully modeled as a mixture of two ancestries: Papuan-related (blue) and Austronesian-related (yellow). The NTT samples require a third ancestry (red-brown) that is associated with Mainland Southeast Asia. For this analysis, the ancient samples from the same site and time period were grouped, as indicated in Table 1. D: results of a qpGraph analysis. This graph indicates that the NM samples have ~30% ancestry from a Papuan-related source (blue) and about ~70% ancestry from an Austronesian-related source (yellow – here the Kankanaey from the Philippines are used as the proxy for this source). In addition to these same two sources, the NTT samples also have ancestry (arrow) from a Mainland Southeast Asian source (red-brown – the Mlabri from Thailand are used as the proxy for this source). Figures and/or data are from [1], which should be consulted for more details.
We first examined the mtDNA and Y chromosome haplogroups, for which one can assign a likely origin as either Austronesian or Papuan, based on the geographic distribution and genetic diversity of the haplogroup [21,22]. Haplogroups assigned as Austronesian are widespread across East and Southeast Asia and have the highest genetic diversity there, and their occurrence in Near Oceania is restricted to coastal New Guinea and offshore islands, precisely where Austronesian-speaking groups are found. Papuan haplogroups are restricted to New Guinea and nearby islands (except for a few which were also carried into Remote Oceania); they occur in both highland and coastal New Guinea and have the highest diversity in the highlands. And the results indicated that several individuals, up to 2.15 kya, had mtDNA and Y chromosome haplogroups with different origins (Table 1), thus providing direct evidence that the Papuan-Austronesian admixture must have begun by at least 2.15 kya (and hence estimates for the admixture date that are less than 2.15 kya must be wrong).
Table 1. Sex and mtDNA and Y chromosome haplogroups for the ancient individuals from Eastern Indonesia. Samples are grouped by age and location. Haplogroups in red are of probable Austronesian origin, and those in blue are of probable Papuan origin. Data are from [1], which should be consulted for more details.
We then carried out the typical analyses of the genome-wide data (obtained by capture-enrichment of ~1.2 million SNPs), beginning with the usual descriptive analyses, namely PCA and ADMIXTURE. Projecting the ancient Indonesian individuals onto a PCA with modern and ancient individuals from Southeast Asia, Indonesia (Western and Eastern), and Near Oceania, showed that the ancient Indonesians fell with modern Eastern Indonesians, as expected. However, the ADMIXTURE results revealed something unexpected: while all samples had both Papuan-related and Austronesian-related ancestry components, all samples from the southern islands of Eastern Indonesia (i.e., the Nusa Tenggaras, NTT) have an additional, Southeast Asian-related (SEA) ancestry component (Figure 3B) that was not found in any of the samples from the northern islands of Eastern Indonesia (i.e., the North Moluccas, NM). The presence of this SEA ancestry component in the NTT samples, but not the NM samples, was confirmed by additional analyses, including qpADM and qpGraph analyses (Figure 3C, 3D).
While it would be of interest to know when the SEA ancestry was contributed to NTT, we could not reliably distinguish between the Austronesian and SEA ancestries in the dating analysis. We therefore focused on dating the Papuan-Austronesian admixture, using a method (DATES; [23]) that can be applied to single, low-coverage ancient genomes. The results (Figure 4) indicate that admixture occurred around 2.5-3 kya, which is close to archaeological and linguistic estimates as to when Austronesians first arrived in the area [15,17].
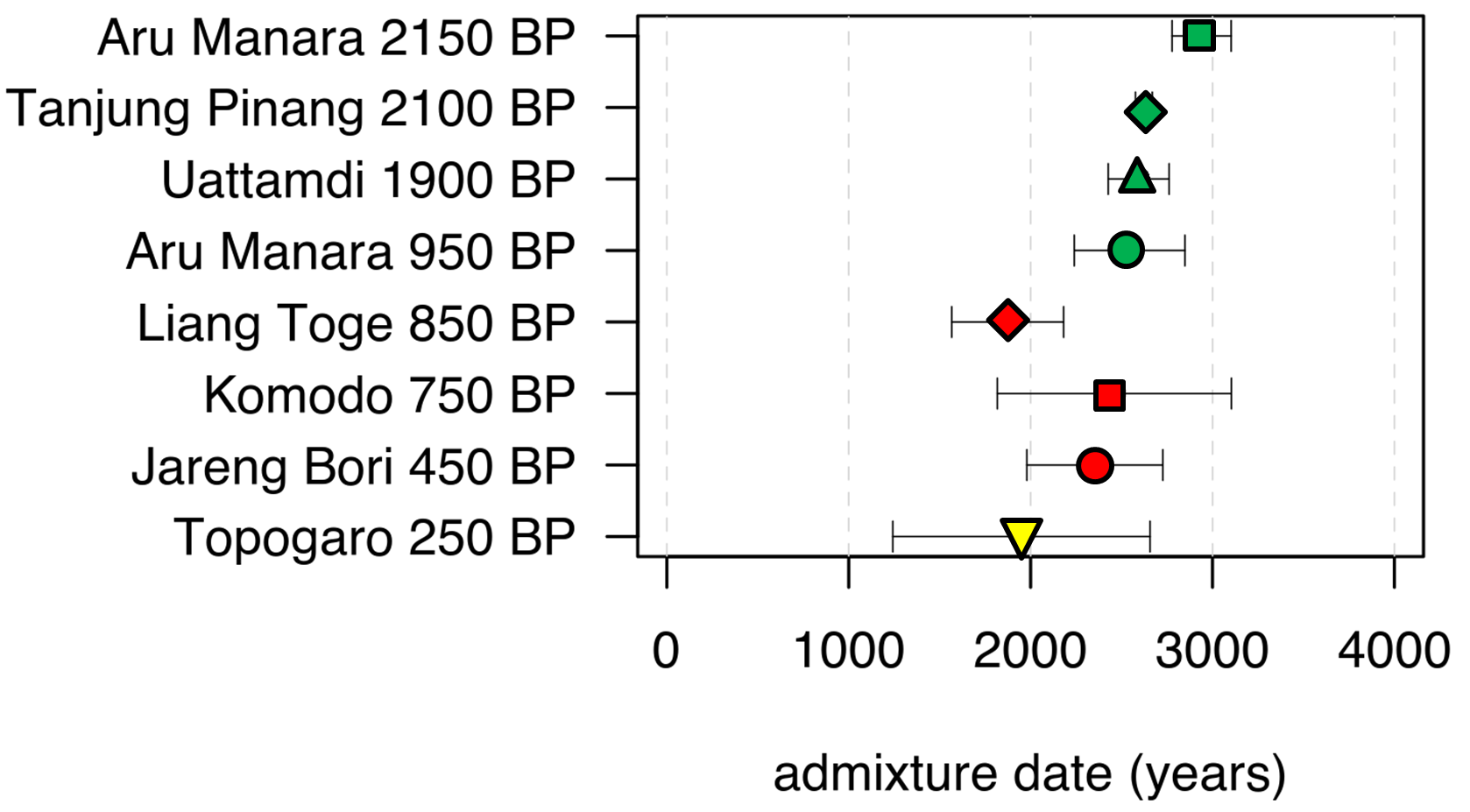
To summarize, the ancient DNA results from Eastern Indonesia indicate that admixture between the incoming Austronesians and the indigenous Papuans probably occurred shortly after the Austronesians arrived in the region. In general, methods for dating admixture events fall into two different categories: those that rely on the length/number of genomic segments of the different ancestries (ancestry blocks: the more numerous and/or shorter the ancestry blocks, the older the admixture event, as over time recombination breaks longer segments into shorter segments); and those that rely on the linkage disequilibrium (LD) that is introduced by admixture (which decays over time). It is notable that the methods that suggested more recent dates for the Papuan-Austronesian admixture (Figure 2) are all LD-based, while the ancestry block methods gave older dates; previous simulation analyses have shown that when there are multiple admixture events over time, LD-based methods tend to give dates that correspond to the most recent event, while ancestry block methods tend to give dates that correspond to the average time of the admixture events [24]. The consistent discrepancy between the dates for the Papuan-Austronesian admixture via LD-based vs. ancestry block methods has two implications: (1) admixture between Austronesians and Papuans did not occur as a single event, but rather as multiple events over time; and (2) the admixture date estimates in Figure 4 are probably under-estimates of the true initial admixture between Austronesians and Papuans. In line with these conclusions, note that older samples tend to give older dates for the ADMIXTURE event (Figure 4), as expected if there have been additional, more recent pulses of admixture.
A further notable finding of this study was the detection of SEA ancestry in the NTT samples, but not in the NM samples. This difference came as a surprise to us, as we were not expecting any significant differences in the history of NTT vs. NM populations. Where this SEA ancestry came from, and when it was contributed to the NTT populations, remain unknown; there is no mention of any such contact in the literature, and none of the archaeologists or linguists we have spoken with see any evidence of such contact. While further work is needed to address from where and when this SEA ancestry arrived, this mystery nicely illustrates the power of ancient DNA analyses to provide novel insights into the history of human populations.
Compared to the settlement of Polynesia, we know much less about the colonization of Micronesia. To quote the noted biological anthropologist Bill Howells [25]: "Many books have been written about where the Polynesians came from but nobody cares a straw about where the Guamanians came from. And yet it is probable that they can tell at least as much about the peopling of the Pacific as can the Polynesians." In fact, in some ways the colonization of Micronesia was even more remarkable than the colonization of Polynesia. For example, archaeological evidence indicates that people were in the Mariana Islands (a chain of 15 islands, of which Guam is the largest and southernmost, stretching across 750 km; Figure 1) by at least 3.5 kya [26], and perhaps as early as 4.3 kya [27]. This is thus around the same time, or perhaps even earlier, than the initial voyages into Polynesia, despite the fact that the Marianas are located over 2000 km from the nearest land (~2200 km from New Guinea and ~2500 km from the Philippines); voyages of comparable length in Polynesia did not occur until nearly 2000 years later. The colonization of the Marianas thus remains one of the more amazing, and yet least known, feats of human exploration.
So where, exactly, did the people who settled the Marianas come from? Chamorro, the indigenous language of Guam, is classified as belonging to the Western Malayo-Polynesian (WMP) subgroup of Austronesian languages; other WMP languages are found exclusively in Western Indonesia (i.e., the islands to the west of Wallace's Line) and the Philippines [28]. Pottery dated to ~3.5 kya in the Marianas shows some similarities to pottery in the Philippines that dates to the same time or even somewhat earlier [29,30]; the linguistic and archaeological evidence would thus suggest an origin from the Philippines or perhaps Western Indonesia. However, the pottery links with the Philippines have been disputed [31], and the language spoken currently in the Marianas may not reflect the language spoken by the first colonizers. Moreover, computer simulations of voyaging that try to take into account seasonal variation in currents and wind and so forth do not find any instances of successful voyaging starting from the Philippines or Western Indonesia [32,33]; instead, successful voyages start from Eastern Indonesia, New Guinea, or the Bismarck Archipelago.
Genetics provides a way to distinguish between settlement from west vs. east of Wallace's Line, as all populations to the west of Wallace's Line lack any Papuan-related ancestry, while all populations to the east of Wallace's Line have significant amounts of Papuan-related ancestry (as we saw in the previous section). So, examining the genetic ancestry of populations from the Marianas for Papuan-related ancestry should provide a relatively simple and straightforward means of distinguishing between these two hypotheses. However, using modern (i.e., contemporary) individuals for this purpose may be problematic, as there could have been changes in ancestry over time that would diminish/erase the ancestry from the initial colonizers. For example, around 1000 years ago stone pillar supports for houses known as latte appear in the Marianas [34], at a time when contact between Pacific islands intensified [12]; moreover, European contact and the associated drastic impacts on native populations began in 1521 with Magellan, and continued with the Manila-Acapulco Spanish galleons that stopped at Guam during voyages that went on from 1565 to 1815. It would thus be preferable to analyze ancient DNA to address this question.
We therefore undertook a study of ancient DNA from the Ritidian Beach Cave site [2] in northern Guam (Figure 5). There were only two individuals from this site, dating to 2.2 kya, so not as old as the original settlement, but older than any subsequent changes in the archaeology that might reflect later colonization. We also could not determine if the two individuals were related or not, as we had insufficient data to answer this question. Still, a sample size of two is bigger than a sample size of zero, so we proceeded anyway with the analysis – the reader should just keep in mind that the results from these two samples may not be representative of the ancient Guam population as a whole.
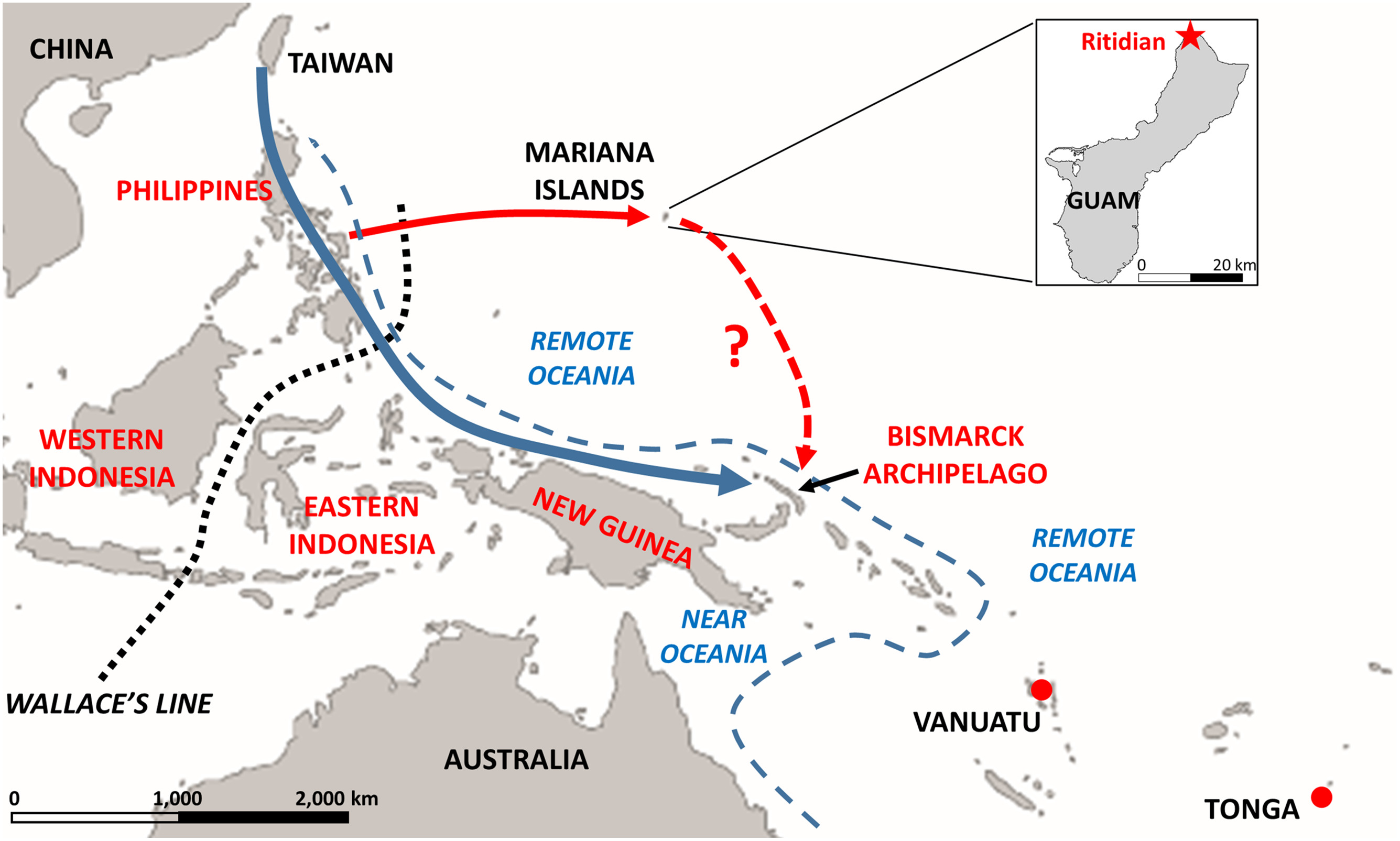
Figure 5. Map showing the location of the Mariana Islands and the Ritidian site on Guam. Also indicated are Wallace's Line and the boundary between Near and Remote Oceania, and the location of the ancient Early Lapita individuals from Vanuatu and Tonga. The blue arrow indicates the conventional view of the migration route from Taiwan to Remote Oceania; the solid red arrow indicates the likely source of the people who colonized the Marianas; and the dashed red arrow with a question mark indicates the highly speculative possibility that the people who went to the Marianas didn't stop there, but continued on and contributed some ancestry to Polynesians. Reprinted with permission from [2].
The usual PCA analysis, with a variety of modern and ancient samples from East Asia, Southeast Asia, and Near and Remote Oceania, shows a gradient of ancestry from East Asia to New Guinea, with the two ancient Guam samples right at the East Asian pole (Figure 6A) – and, intriguingly, close to early Lapita samples from Vanuatu and Tonga, in Polynesia (more on this later). So, this analysis suggests no Papuan-related ancestry in the ancient Guam samples. We investigated this in more detail by a variety of analyses, including outgroup f3 and f4 statistics. The f3 analyses indicate how much genetic drift is shared between the ancient Guam samples and a variety of other populations, and the results show that the ancient Guam samples are closest (i.e., share the most genetic drift) with the early Lapita samples, an ancient sample from the Philippines, and modern samples from the Philippines and Taiwan (Figure 6B). They share less genetic drift with Papuans than with any other population in the analysis (except Europeans). The f4 analyses test if a group shares more ancestry with New Guinea highlanders than do the Kankanaey, a modern group from the Philippines which does not show any indication of Papuan-related ancestry. All groups tested from Oceania show significant amounts of Papuan-related ancestry – except for the two ancient Guam samples (Figure 6C). None of the analyses we did detected any indication of any Papuan-related ancestry in the ancient Guam samples, and we consistently found that samples from the Philippines were genetically close to the ancient Guam samples (so, it would appear that we need to rethink the computer simulations of ancient voyaging!). A later study that included many more samples from Guam came to the same conclusion of a likely origin from the Philippines [35].
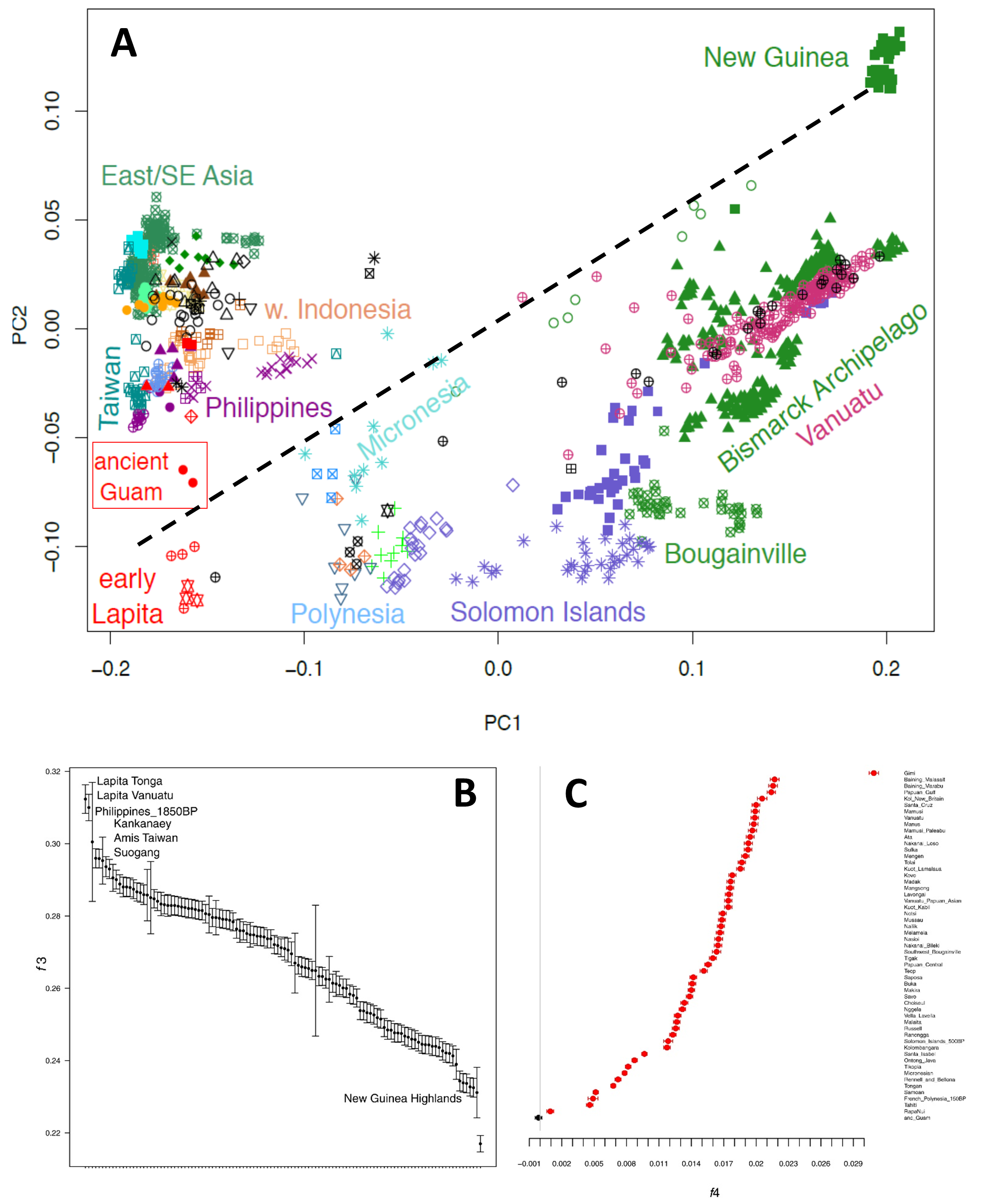
Figure 6. Results of analyses of the ancient Guam samples. A: plot of the first two principal components from the PCA. The dashed line indicates the gradient of ancestry from purely Asian (bottom left) to purely Papuan (top right); note that the ancient Guam samples are located near other samples that completely lack Papuan ancestry (and are close to the Early Lapita samples). B: Results of an outgroup f3 analysis, indicating that the ancient Guam samples are most similar to (i.e., share the most drift with) the Early Lapita samples and ancient and modern samples from the Philippines and Taiwan; they are most distant from Papuans (New Guinea Highlanders). C: results of an f4 analysis of the form (Test, Kankanaey; New Guinea Highlanders, Mbuti Pygmy), where "Test" is each population on the right of the plot. Significant positive values (in red) indicate that the Test population shares excess ancestry with New Guinea Highlanders compared to the Kankanaey from the Philippines, while non-significant values (in black) indicate no excess Papuan ancestry in the test population compared to the Kankanaey. The populations included in the analysis in the "Test" position are a variety of Near and Remote Oceanian populations, and all of them give significant positive results, indicating that all of them have more Papuan related ancestry than do the Kankanaey – except for the ancient Guam samples, at the bottom of the plot. Figures and/or data are from [2], which should be consulted for more details.
We also found some shared ancestry between the ancient Guam samples and the early Lapita samples from Polynesia. In principle, this could reflect that the ancient settlers of Guam and of Polynesia came from the same (or closely-related) source populations in the Philippines. However, it is intriguing to speculate that perhaps the people who first went to the Marianas didn't simply stop there, but instead their descendants continued voyaging and perhaps even contributed some ancestry to Polynesians (Figure 5). Further research is needed to investigate this suggestion; for now, I leave the final word on this topic to Bill Howells [25]: "What about a Micronesian route [for the colonization of Polynesia]? It is not in favor with the anthropologists, though after all it was not anthropologists who settled Polynesia."
These two studies nicely illustrate the power of ancient DNA studies to address long-standing controversies concerning when Austronesians began admixing with indigenous Papuan groups in Eastern Indonesia, and where the initial colonizers of the Marianas came from. They also illustrate how ancient DNA can provide novel insights, such as the SEA ancestry in the NTT of Eastern Indonesia, and the close relationship between the early settlers of Guam and those of Polynesia. And there are certainly other studies one can point to, such as the recent study of pre-colonial individuals from Rapanui [36], which provides definitive proof of native American ancestry in Polynesia prior to the arrival of Europeans. Another nice feature of this study is that the ancient DNA indicates that there was never any population collapse due to over-exploitation of resources on the island, as has been commonly supposed [37]. Still, ancient DNA studies in this (and many other) parts of the world are limited by issues relating to DNA preservation: DNA simply does not preserve well in hot/humid conditions. While people will certainly keep trying, for every success there are many failures, and so there will continue to be a role for studies of modern populations, and for trying to extract as much information as we can from modern genomes from Oceania [38-41].
Not applicable.
Not applicable.
Not applicable.
The original research described in these two case studies was funded by the Max Planck Society.
Mark Stoneking is a member of the Editorial Board of the journal Human Population Genetics and Genomics. The author was not involved in the journal’s review of or decisions related to this manuscript. The author has declared that no other competing interests exist.
I thank the organizers of the Wellcome Trust meeting for the opportunity to present this work, and the editors of HPGG for the invitation to contribute to the special issue. I also thank my co-authors on these two studies, especially the archaeologists who provided samples and shared their knowledge about this interesting part of the world, and the students and postdocs who actually did the work presented here. Funding for these two studies was provided by the Max Planck Society.
| 1. | Oliveira S, Nägele K, Carlhoff S, Pugach I, Koesbardiati T, Hübner A, et al. Ancient genomes from the last three millennia support multiple human dispersals into Wallacea. Nat Ecol Evol. 2022;6(7):1024-1034. [Google Scholar] [CrossRef] |
| 2. | Pugach I, Hubner A, Hung HC, Meyer M, Carson MT, Stoneking M. Ancient DNA from Guam and the peopling of the Pacific. Proc Natl Acad Sci U S A. 2021;118(1):e2022112118. [Google Scholar] [CrossRef] |
| 3. | Matisoo-Smith E, Gosling AL. Understanding the human settlement of the Pacific – Are we there yet? J Archaeol Sci. 2025;180:106307. [Google Scholar] [CrossRef] |
| 4. | Stoneking M. Insights from ancient human DNA into the colonization of Oceania. In: Elias S, editor. Encyclopedia of Quaternary Science. 3rd ed. UK: Elsevier; 2025. p. 742-750. [CrossRef] |
| 5. | Stoneking M. An Introduction to Molecular Anthropology. 2nd ed. Hoboken, New Jersey: John Wiley and Sons, Incorporated; 2025. [Google Scholar] |
| 6. | O'Connell JF, Allen J, Williams MAJ, Williams AN, Turney CSM, Spooner NA, et al. When did Homo sapiens first reach Southeast Asia and Sahul? Proc Natl Acad Sci U S A. 2018;115(34):8482-8490. [Google Scholar] [CrossRef] |
| 7. | Kirch PV. Peopling of the Pacific: A Holistic Anthropological Perspective. Annu Rev Anthropol. 2010;39(1):131-148. [Google Scholar] [CrossRef] |
| 8. | Sear DA, Allen MS, Hassall JD, Maloney AE, Langdon PG, Morrison AE, et al. Human settlement of East Polynesia earlier, incremental, and coincident with prolonged South Pacific drought. Proc Natl Acad Sci U S A. 2020;117(16):8813-8819. [Google Scholar] [CrossRef] |
| 9. | Gaffney D, Tanudirjo DA, Djami ENI, Mas'ud Z, Macap AR, Russell T, et al. Human dispersal and plant processing in the Pacific 55 000–50 000 years ago. Antiquity. 2024;98(400):885-904. [Google Scholar] [CrossRef] |
| 10. | Mondal M, Andre M, Pathak AK, Brucato N, Ricaut FX, Metspalu M, et al. Resolving out of Africa event for Papua New Guinean population using neural network. Nat Commun. 2025;16(1):6345. [Google Scholar] [CrossRef] |
| 11. | Bellwood P. Prehistory of the Indo-Malaysian Archipelago. Canberra, Australia: ANU Press; 2007. [Google Scholar] [CrossRef] |
| 12. | Kirch PV. On the Road of the Winds: an Archaeological History of the Pacific Islands Before European Contact. Oakland, CA: University of California Press; 2017. [Google Scholar] |
| 13. | Crowley T. Austronesian languages. In: Brown K, Ogilvie S, editors. Concise Encyclopedia of Languages of the World. Oxford, UK: Elsevier; 2006. p. 96-105. |
| 14. | Serva M, Petroni F, Volchenkov D, Wichmann S. Malagasy dialects and the peopling of Madagascar. J R Soc Interface. 2012;9(66):54-67. [Google Scholar] [CrossRef] |
| 15. | Gray RD, Drummond AJ, Greenhill SJ. Language Phylogenies Reveal Expansion Pulses and Pauses in Pacific Settlement. Science. 2009;323(5913):479-483. [Google Scholar] [CrossRef] |
| 16. | Pawley A. Papuan languages. In: Brown K, Ovilgie S, editors. Concise Encyclopedia of Languages of the World. Oxford, UK: Elsevier; 2006. p. 836-844. |
| 17. | Bellwood P. The First Farmers: The Origins of Agricultural Societies. Oxford: Blackwell; 2005. [Google Scholar] |
| 18. | Hudjashov G, Karafet TM, Lawson DJ, Downey S, Savina O, Sudoyo H, et al. Complex Patterns of Admixture across the Indonesian Archipelago. Mol Biol Evol. 2017;34(10):2439-2452. [Google Scholar] [CrossRef] |
| 19. | Lipson M, Loh P-R, Patterson N, Moorjani P, Ko Y-C, Stoneking M, et al. Reconstructing Austronesian population history in Island Southeast Asia. Nat Commun. 2014;5(1):4689. [Google Scholar] [CrossRef] |
| 20. | Xu S, Pugach I, Stoneking M, Kayser M, Jin L. Genetic dating indicates that the Asian–Papuan admixture through Eastern Indonesia corresponds to the Austronesian expansion. Proc Natl Acad Sci U S A. 2012;109(12):4574-4579. [Google Scholar] [CrossRef] |
| 21. | Delfin F, Myles S, Choi Y, Hughes D, Illek R, van Oven M, et al. Bridging Near and Remote Oceania: mtDNA and NRY Variation in the Solomon Islands. Mol Biol Evol. 2011;29(2):545-564. [Google Scholar] [CrossRef] |
| 22. | Kayser M, Brauer S, Cordaux R, Casto A, Lao O, Zhivotovsky LA, et al. Melanesian and asian origins of Polynesians: mtDNA and Y chromosome gradients across the Pacific. Mol Biol Evol. 2006;23(11):2234-2244. [Google Scholar] [CrossRef] |
| 23. | Narasimhan VM, Patterson N, Moorjani P, Rohland N, Bernardos R, Mallick S, et al. The formation of human populations in South and Central Asia. Science. 2019;365(6457):eaat7487. [Google Scholar] [CrossRef] |
| 24. | Pugach I, Duggan AT, Merriwether DA, Friedlaender FR, Friedlaender JS, Stoneking M. The gateway from Near into Remote Oceania: new insights from genome-wide data. Mol Biol Evol. 2018;35(4):871-886. [Google Scholar] [CrossRef] |
| 25. | Howells WW. The Pacific Islanders. New York: Charles Scribner's Sons; 1973. [Google Scholar] |
| 26. | Carson MT. Peopling of Oceania: clarifying an initial settlement horizon in the Mariana Islands at 1500 BC Radiocarbon. 2020;62(6):1733-1754. [CrossRef] |
| 27. | Athens JS, Dega MF, Ward JV. Austronesian colonization of the Mariana Islands: the palaeoenvironmental evidence. Bulletin Indo-Pacific Prehistory Association. 2004;24:21-30. [Google Scholar] |
| 28. | Blust R. The Austronesian Languages. Canberra, Australia: Asia-Pacific Linguistics, ANU; 2013. [Google Scholar] |
| 29. | Carson MT, Hung HC, Summerhayes G, Bellwood P. The Pottery Trail From Southeast Asia to Remote Oceania. J Island Coastal Archaeol. 2013;8(1):17-36. [Google Scholar] [CrossRef] |
| 30. | Hung HC, Carson MT, Bellwood P, Campos FZ, Piper PJ, Dizon E, et al. The first settlement of Remote Oceania: the Philippines to the Marianas. Antiquity. 2011;85(329):909-926. [Google Scholar] [CrossRef] |
| 31. | Winter O, Clark G, Anderson A, Lindahl A. Austronesian sailing to the northern Marianas, a comment on Hung et al. (2011). Antiquity. 2012;86(333):898-910. [Google Scholar] [CrossRef] |
| 32. | Fitzpatrick SM, Callaghan RT. Estimating trajectories of colonisation to the Mariana Islands, western Pacific. Antiquity. 2013;87(337):840-853. [Google Scholar] [CrossRef] |
| 33. | Montenegro A, Callaghan RT, Fitzpatrick SM. Using seafaring simulations and shortest-hop trajectories to model the prehistoric colonization of Remote Oceania. Proc Natl Acad Sci U S A. 2016;113(45):12685-12690. [Google Scholar] [CrossRef] |
| 34. | Carson MT. An overview of latte period archaeology. Micronesica. 2012;42:1-79. [Google Scholar] |
| 35. | Liu YC, Hunter-Anderson R, Cheronet O, Eakin J, Camacho F, Pietrusewsky M, et al. Ancient DNA reveals five streams of migration into Micronesia and matrilocality in early Pacific seafarers. Science. 2022;377(6601):72-79. [Google Scholar] [CrossRef] |
| 36. | Moreno-Mayar JV, Sousa da Mota B, Higham T, Klemm S, Gorman Edmunds M, Stenderup J, et al. Ancient Rapanui genomes reveal resilience and pre-European contact with the Americas. Nature. 2024;633(8029):389-397. [Google Scholar] [CrossRef] |
| 37. | Diamond J. Collapse: How Societies Choose to Fail or Succeed. New York City, New York: Penguin Random House; 2005. [Google Scholar] |
| 38. | Choin J, Mendoza-Revilla J, Arauna LR, Cuadros-Espinoza S, Cassar O, Larena M, et al. Genomic insights into population history and biological adaptation in Oceania. Nature. 2021;592(7855):583-589. [Google Scholar] [CrossRef] |
| 39. | Ioannidis AG, Blanco-Portillo J, Sandoval K, Hagelberg E, Barberena-Jonas C, Hill AVS, et al. Paths and timings of the peopling of Polynesia inferred from genomic networks. Nature. 2021;597(7877):522-526. [Google Scholar] [CrossRef] |
| 40. | Ioannidis AG, Blanco-Portillo J, Sandoval K, Hagelberg E, Miquel-Poblete JF, Moreno-Mayar JV, et al. Native American gene flow into Polynesia predating Easter Island settlement. Nature. 2020;583(7817):572-577. [Google Scholar] [CrossRef] |
| 41. | Jacobs GS, Hudjashov G, Saag L, Kusuma P, Darusallam CC, Lawson DJ, et al. Multiple deeply divergent Denisovan ancestries in Papuans. Cell. 2019;177(4):1010-1021. [Google Scholar] [CrossRef] |

![]()
Copyright © 2026 Pivot Science Publications Corp. - unless otherwise stated | Terms and Conditions | Privacy Policy




