1. Introduction
Single-cell analysis has emerged as a cornerstone of modern biological research, offering unparalleled insights into the nuanced complexities of cellular behavior and heterogeneity [1-6]. In contrast to conventional population-based approaches, which aggregate data from numerous cells, single-cell analysis looks into the unique characteristics of individual cells, providing a comprehensive view of cellular diversity. This level of scrutiny allows researchers to uncover subtle yet significant variations in cellular states, responses to environmental cues, and molecular signatures that may go unnoticed in bulk analyses [7, 8]. Such granularity is indispensable for deciphering complex biological processes, including embryonic development, disease pathogenesis, and therapeutic interventions [9-12]. By dissecting the cellular landscape at the single-cell level, scientists can unlock a deeper understanding of cellular function and behavior, paving the way for transformative discoveries in biomedical science [13].
Cancer is featured with the presence of heterogeneous cell populations within tumors, exhibiting variations in gene expression, phenotype, and functional properties [14-18]. This heterogeneity arises from genetic mutations, epigenetic modifications, and microenvironmental influences, contributing to tumor evolution, metastasis, and therapy resistance [19-23]. Understanding cell heterogeneity in cancer is crucial for several objectives [1]. Treatment response and resistance: heterogeneous cell populations within tumors respond differently to therapies, leading to variable treatment outcomes [14,24-30]. Subpopulations of cells with intrinsic or acquired resistance mechanisms can evade cytotoxic effects, leading to treatment failure and disease relapse. By characterizing and targeting specific cell subsets, clinicians can devise more effective treatment strategies tailored to individual patients, improving therapeutic outcomes [2]. Disease progression and metastasis: cell heterogeneity plays a critical role in cancer progression and metastasis [6,31-33]. Subclones with enhanced migratory and invasive properties can disseminate from primary tumors, colonize distant sites, and establish metastatic lesions. The presence of distinct cell populations within tumors contributes to the formation of heterogeneous metastases, complicating treatment and prognosis. Understanding the cellular dynamics driving metastatic spread is essential for developing interventions to prevent or inhibit metastasis [3]. Biomarker discovery and patient stratification: cell heterogeneity offers valuable insights into disease biology and patient stratification [34-40]. Identification of specific cell populations associated with aggressive phenotypes, poor prognosis, or treatment resistance can inform the development of prognostic biomarkers and therapeutic targets. By stratifying patients based on their molecular and cellular profiles, clinicians can personalize treatment regimens, optimize therapeutic efficacy, and minimize adverse effects [4]. Precision Medicine and Therapeutic Innovation: In the era of precision medicine, targeting the unique molecular and cellular features of individual tumors is paramount [41-47]. Single-cell analysis enables the identification of rare cell populations, characterization of signaling pathways, and assessment of therapeutic vulnerabilities. By leveraging this information, researchers can design precision therapies that selectively target tumor cells while sparing normal tissues, enhancing treatment specificity, and minimizing off-target effects.
Considering the important role of cellular heterogeneity in cancer, elucidating the relationship between phenotype and molecular genotype is essential. Phenotype, often representing traits like drug resistance, invasiveness, or responsiveness to stimuli, directly informs targeted therapeutic interventions. Conversely, genotype provides the molecular framework governing these phenotypic traits. Notably, a single phenotype can be influenced by multiple genotypic characteristics, and genotypic alterations can result in diverse phenotypic outcomes depending on contextual factors. These sophisticated interplays underscore the significance of correlating phenotypic and genotypic domains at single-cell resolution, offering invaluable insights into the mechanisms of tumor development, progression, resistance, and metastasis. Understanding the genotype-phenotype relationship also aids in discovering novel therapeutic targets. By elucidating how specific genetic changes translate into oncogenic phenotypes, researchers can identify new pathways and genes to target with innovative drugs, thus expanding the arsenal of therapeutic options available to treat various cancers.
To identify cells exhibiting distinct phenotypes for subsequent molecular genotypic analysis, Fluorescence-Activated Cell Sorting (FACS) and Magnetic-Activated Cell Sorting (MACS) apparatuses are the first-generation approach, enabling cell sorting based on surface markers or enzymatic activity. These methodologies offer a high throughput and are prevalent in research institutes, with FACS instruments becoming increasingly adept at sorting cells using a growing array of fluorescent markers. Nonetheless, there are significant limitations. Firstly, sorting relies on a single snapshot measurement, lacking the capacity to monitor temporal processes within individual cells. Moreover, FACS typically sorts cells post-trypsinization, resulting in the loss of original morphological features, spatial context, and complicating the differentiation based on shape or subcellular organelle distribution. Additionally, FACS fails to segregate cells according to functional behaviors, such as motility or persistence. In contrast, microscopy-based cell tracking offers a broader array of functionalities, encompassing cell behavior exploration, biosensor kinetics, enzyme reaction kinetics, gene expression dynamics, and subcellular localization assessments. However, although microscopy enables phenotypic observation, there remains a necessity to selectively isolate cells of interest for subsequent genotypic analysis to decipher the underlying mechanisms governing phenotype or morphology. In this review, we will elucidate innovative strategies for selectively isolating cells based on targeted phenotypes, including selective cell detachment and retrieval, on-site cell marking for enrichment in subsequent stages, and the droplet-based approach, which facilitates observation and sorting.
2. Single-cell Labeling and Isolation Methods
2.1 Selective cell retrieval by robotic aspiration or laser detachment
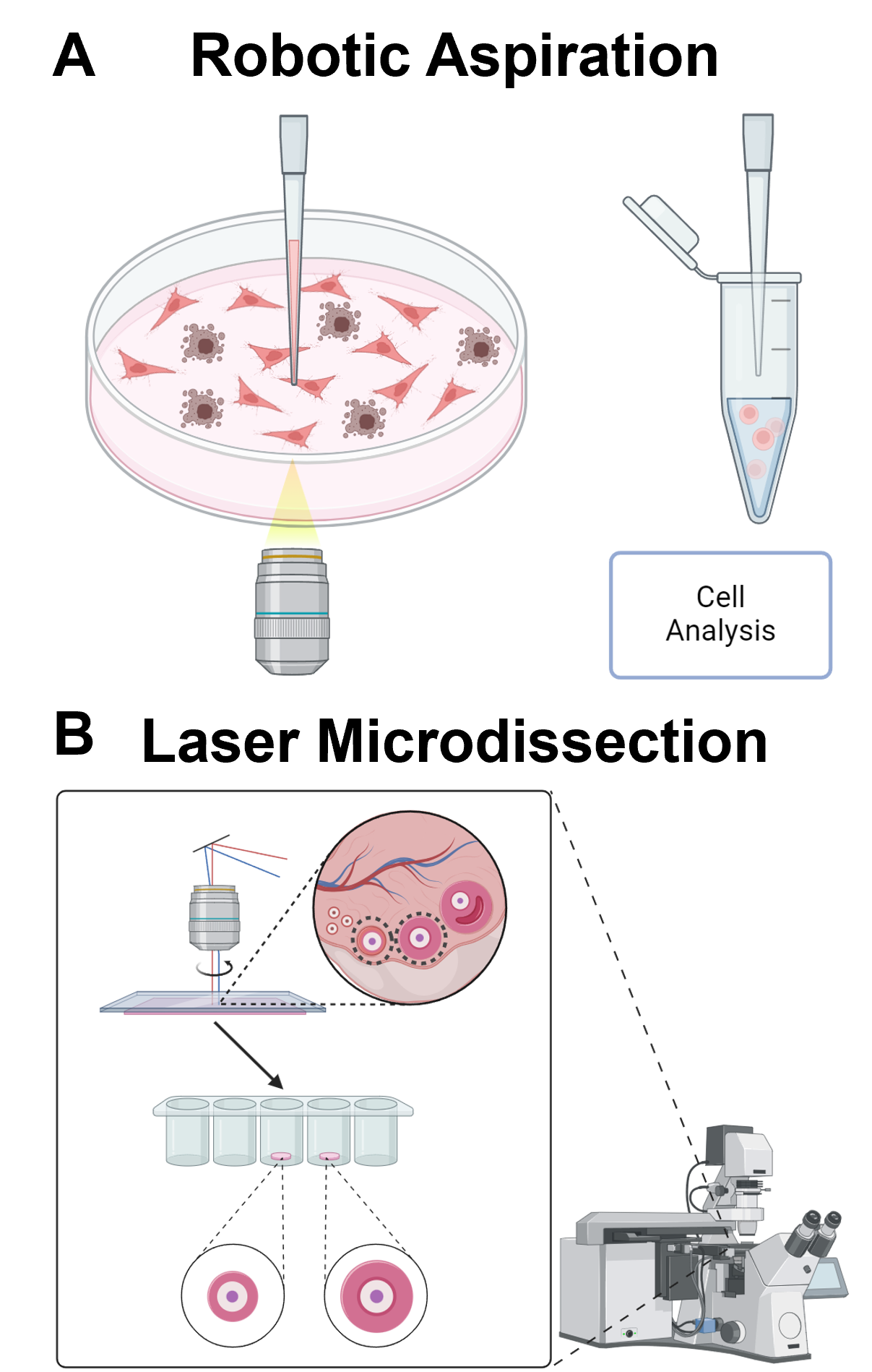
For analyzing target cells with desired phenotype, a straightforward idea is to selectively retrieve them for further downstream analysis. Robotic aspiration and laser microdissection are two conventional strategies. The robotic aspiration technique for single-cell retrieval leverages advanced micromanipulation technologies integrated with computer vision and motion control algorithms to achieve precise and efficient handling of individual cells (Figure 1A) [48-50]. The core principle involves a pick-and-place system that can visually track a cell in real-time and accurately control positioning devices to pick up, transfer, and deposit a single cell at a specific location on a desired substrate. This process starts with a traditional glass micropipette, which is used to aspirate cells either partially or wholly. The technique incorporates computer vision to identify and track a target cell within the culture environment. Once the target cell is identified, the motion control algorithms coordinate the movement of the micropipette to approach and aspirate the cell. After aspiration, the robotic system precisely transfers the cell to a predetermined location on the substrate and deposits it for downstream analysis.
Laser capture microdissection (LCM) is a conventional precise technique used to isolate specific cells or regions from heterogeneous tissue samples by employing a focused laser beam to cut around and capture the cells of interest (Figure 1B) [51-52]. The process involves preparing thin tissue sections or cell cultures on special slides, visualizing the sample under a microscope to identify target cells, and using the laser to either melt an adhesive film or directly cut around the cells for retrieval. There are also variations of laser cell extraction, including the microcapillary single cell analysis and laser extraction (μSCALE) system, which disrupts the surface tension of a microcapillary, ejecting its contents onto an cell isolation substrate with a UV pulsed laser [53]. Both aspiration and LCM offers several advantages, including availability of commercial instrument, high precision, compatibility with downstream DNA, RNA, and protein analyses, and minimal sample processing. However, these approaches also have drawbacks, such as high initial cost of purchasing the instrument, time-consuming manual identification and capture, potential risk of sample damage from aspiration or laser, and a limitation to thin tissue sections, which may not represent the tumor microenvironment. Despite these challenges, robotic aspiration and LCM remain useful and reliable strategies for isolating specific cells with minimal contamination, enabling detailed molecular profiling and analysis.
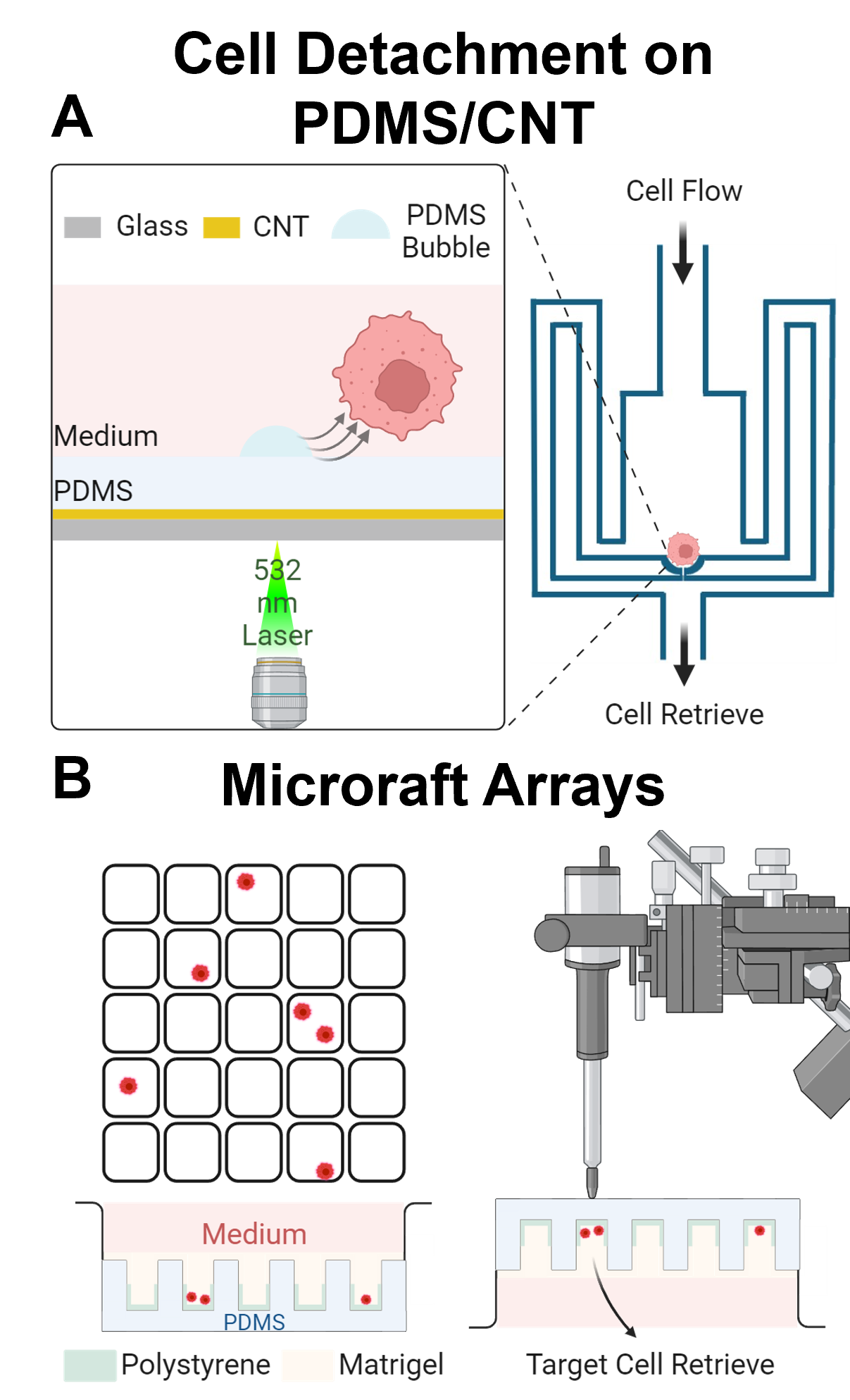
Another laser-based strategy for retrieving cells from enclosed microfluidics involves a multi-functional bottom substrate composed of a carbon nanotube-polydimethylsiloxane (CNT-PDMS) composite film (Figure 2A) [54]. In this microfluidic system, once target cells are identified, a nanosecond laser pulse is applied to the CNT-PDMS film to generate micro-bubbles. The carbon nanotubes (CNTs) within the PDMS matrix convert the incident optical energy into thermal energy, exploiting the composite's inherent nanoscale porosity to expand and merge nano-bubbles [55,56]. This process results in rapid generation of heat-induced bubbles from within the PDMS, which then burst through the film surface. The targeted cell experiences shear forces from the bubble rupture and lateral displacement caused by bubble expansion, effectively detaching it from the surface. The CNT-PDMS composite film offers several advantages: (1) it supports highly biocompatible cell culture, (2) it can be easily integrated into microfluidic fabrication processes, and (3) it enables the generation of optically driven shear forces (or micro-bubbles) that disrupt cell adhesion to the surface. Additionally, the low thermal conductivity of PDMS protects cells from the heat generated in the CNT layer, thereby preserving cell viability. While this method is innovative and well-integrated with microfluidic systems, it requires a specific nanosecond pulse laser source, and a commercial system for this technology is not yet available.
2.2 Microraft array for single-cell or clonal isolation
Microraft arrays (MRAs) represent another engineering solution for selectively single cell or clonal retrieval (Figure 2B) [57,58]. It involves a substrate patterned with thousands of tiny, detachable polymeric "rafts," each capable of capturing and supporting individual cells. After cells are seeded onto the array and allowed to grow in isolation, they can be identified and analyzed using microscopy. The cells with target phenotype are then retrieved by dislodging the specific rafts using a robotic micromanipulator or a microfabricated needle, which can be assisted with laser cutting. This gentle, non-destructive process preserves cell viability and allows for detailed downstream analysis, such as single-cell sequencing. The microraft array is versatile, supporting various cell types, though it requires specialized microraft arrays or expertise for fabrication and manipulation. In addition, it is difficult to separate two daughter cells from one mother cell, as both daughter cells will be on the same microraft. Despite some limitations, this technology is reliable and valuable in minimizing effects on cells.
2.3 Optical traps for selective single-cell isolation based on phenotypes
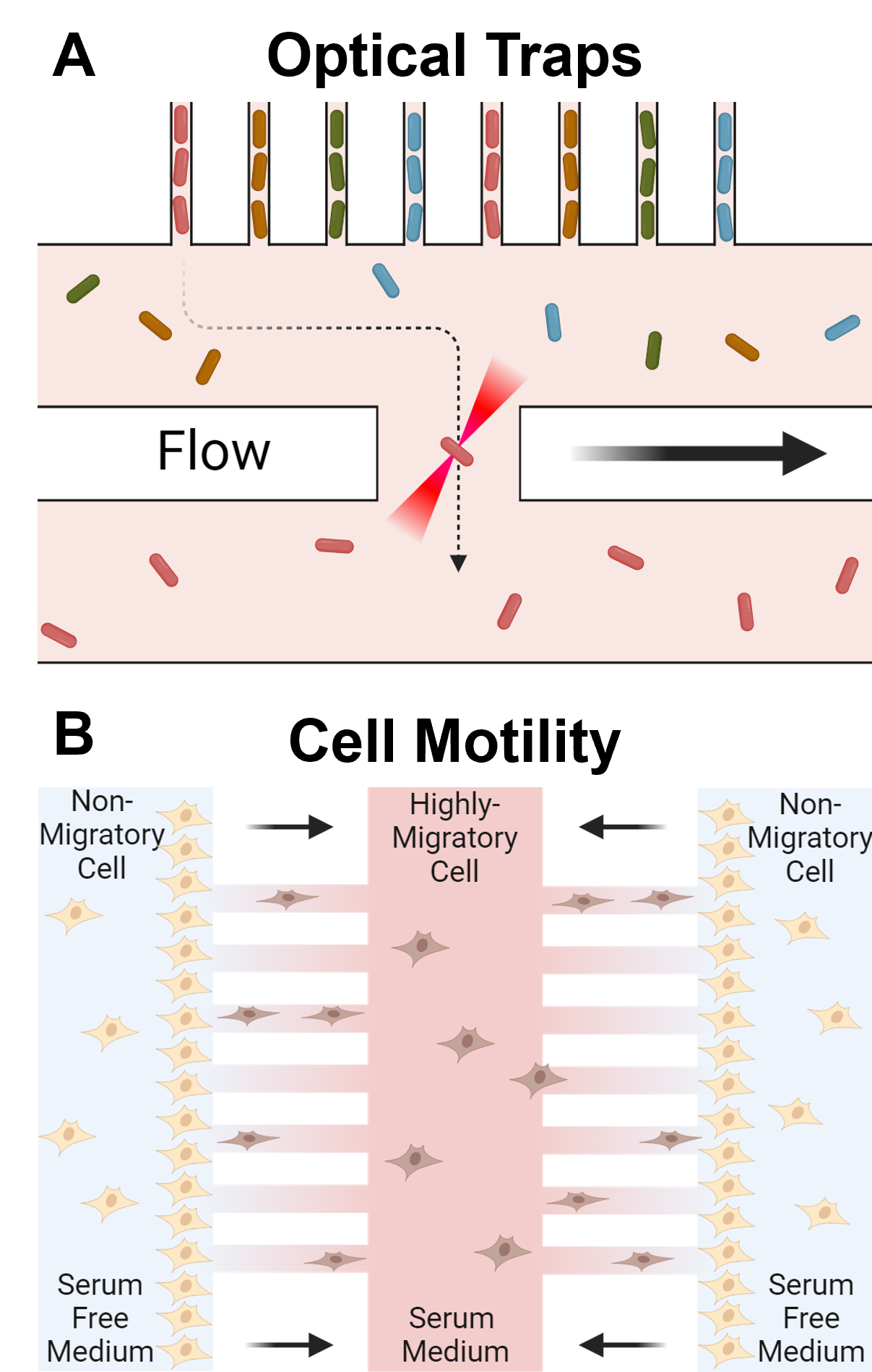
Optical traps, also known as optical tweezers, are another sophisticated tool that uses highly focused laser beams to manipulate and isolate single cells with high precision (Figure 3A) [59-63]. The core principle involves the gradient force exerted by the laser light on the cell, which allows for non-contact manipulation and detailed measurement of cellular properties. To build an optical trap, a laser beam is tightly focused through a microscope objective lens, creating a highly intense electric field at the focal point. The intensity is critical as it generates a gradient force that draws the cell toward the region of highest light intensity, effectively trapping it. Concurrently, a scattering force from the momentum transfer of photons acts in the direction of light propagation. For stable trapping, the gradient force must surpass the scattering force, which is accomplished through precise laser focusing. Once the cell is stably trapped, it can be manipulated by moving the laser beam or adjusting the focus, facilitating various types of single-cell phenotypic analyses. These analyses include measuring the mechanical properties of the cell by applying known forces and observing deformation, thereby assessing stiffness and elasticity. Optical tweezers can also be used for force spectroscopy to measure forces involved in cellular processes like division or migration. Additionally, intracellular transport can be studied by tracking the movement of organelles or vesicles within the trapped cell, providing insights into cellular dynamics. In addition to inducing lateral cell movement, optical tweezers can be employed to exert forces in the vertical direction, enabling the selective levitation of single cells from microwells for isolation [64]. The technique is also valuable for single-molecule studies, where specific molecules are attached to the cell's surface or interior to investigate molecular interactions and dynamics within the cell [65]. Optical tweezers facilitate non-invasive observation and manipulation of cells, preserving their viability and enabling real-time analysis of dynamic processes in their native environment. While this method is effective for non-adherent objects, such as yeast, bacteria, and immune cells, it is not suitable for moving adherent cancer cells. Additionally, the technique has limitations, including the complexity of the optical setup and moderate to low throughput for cell sorting.
2.4 Selective cell isolation based on cellular motility or deformability
Under specific conditions, certain phenotypes can facilitate selective cell isolation. For instance, cell separation based on motility and chemotaxis, critical in cancer dissemination [66-70], can be achieved using engineered microfluidic devices [5,6,71,72]. These devices typically feature paired wells connected by cell migration channels (Figure 3B). Initially, cells are seeded into one well. After allowing for cell adhesion to the substrate, a chemoattractant is introduced into the opposite well to induce cell migration through the channels. The fast-moving cells that migrate to the other well can then be easily harvested through trypsinization. Similarly, cell deformability, a phenotype associated with cancer invasion, can be exploited for separation using a microfluidic device [73-74]. A microfluidic mechanical deformability chip, for example, employs artificial microbarriers to differentiate flexible cells from stiff ones using hydrodynamic forces [75-76]. More elastic cells pass through the microbarriers and exit the separation chip, while stiffer cells remain trapped. Although those microfluidic platforms are highly effective for phenotypic-based cell separation, their applicability is limited to phenotypes characterized by distinct motility or deformability, thus restricting its applicability to a broader range of applications.
2.5 On-site cell marking for further separation
In addition to retrieving cells immediately, an alternative is to mark cells with certain phenotype with fluorescence or other markers. The marked cells can be separated in the following steps [77-82]. SPOTlight is a representative example of this concept (Figure 4A) [83]. Target cells, such as bacteria, yeast, or mammalian cells, can be made optically taggable by introducing photo-transformable fluorescent proteins or dyes that shift from a dim to bright state or change colors when excited by specific light wavelengths. This tagging can be applied to naturally heterogeneous cell populations as well as cell libraries, such as those used in siRNA screens. Cells are imaged to identify phenotypes of interest, which are quantified at single-cell resolution. Cells of interest are selected based on their phenotypic profiles and optically tagged via single-cell illumination. The tagged cells are then identified and isolated using FACS. Subsequently, the selected cells undergo genotyping or further characterization as needed. Alternatively, cells can be labeled by attaching streptavidin-coated magnetic beads to their membranes using the lasers of a confocal microscope for sorting [84,85]. A simple magnet can then be used to achieve highly specific isolation of the labeled cells, which remain healthy and continue to proliferate normally. The advantages of optical tagging include its high precision, allowing cellular resolution targeting; temporal control, enabling the study of rapid biological events; versatility across various cell types and applications, including optogenetic control, photothermal therapy, and fluorescence activation; and real-time feedback, enhancing accuracy and efficiency. However, the use of phototransformable fluorescent proteins or dyes and the need for prolonged or intense single-cell illumination could potentially affect cell viability or alter cell status. Additionally, the throughput of this method might be constrained by the time required for illumination, image collection, and processing to select cells exhibiting the desired phenotypes. If the proportion of labeled cells is relatively low, significant cell loss may occur during the subsequent FACS step.
2.6 Single-cell phenotypic analysis and sorting within droplets
Another promising approach involves performing single-cell phenotypic assays within droplets, which can be efficiently sorted using microfluidics or FACS. Droplet-based microfluidics entails creating and manipulating minute droplets within microchannels, each acting as isolated microreactors [86-88]. This technology facilitates high-throughput single-cell analysis, biochemical reactions, and assays by compartmentalizing samples, providing precise control over the microenvironment, and enabling efficient, scalable biological and chemical experimentation. For example, droplet-based microfluidics can isolate individual antibody-secreting cells from a large number of non-secreting cells at high throughput [89-91]. These principles and methods are highly adaptable for various applications. By screening for antigen binding, the droplet system rapidly selects cells that secrete target-specific antibodies. Its high throughput and ability to preserve cell viability enable the testing of primary blood cells from patients without the need for cell immortalization. The fluorescence-based binding assay can be adapted to measure any secreted molecule with a fluorescently labeled ligand, such as insulin, chemokines, or growth factors. Additionally, using fluorogenic substrates, the protocol can be tailored for high-throughput screening, directed enzyme evolution, or screening antibodies and molecules that inhibit enzyme activity. This versatile approach can analyze a wide range of cells, including bacteria, yeast, and mammalian cells, by examining the activities of secreted, cell-surface, or intracellular proteins. However, it is important to note that not all assays are well-suited for the droplet format. Although droplet splitting and fusion allow operations such as washing or adding new reagents, completely removing reagents from an initial reaction before introducing new ones is challenging [92-94]. Furthermore, adhesion-dependent epithelial cancer cells might suffer from anoikis due to loss of anchorage [29,95,96], making long-term culture of adherent cancer cells within droplets difficult and limiting the applications in cancer research.
Building on the concept of cell phenotypic assays in droplets, hydrogel nanovials have been developed for single-cell phenotypic analysis and sorting (Figure 4B) [97-99]. The core principle involves encapsulating individual cells within hydrogel nanovials, which are microscopic, suspendable containers made of biocompatible hydrogel material. These nanovials are engineered with functionalized surfaces to capture and retain single cells along with reagents necessary for specific assays. Each hydrogel nanovial provides an isolated microenvironment where individual cells can undergo various functional assays, such as enzymatic activity, protein secretion, or cell-cell interactions. The hydrogel material allows for the diffusion of small molecules and reagents, facilitating real-time monitoring of cellular responses. The nanovials can be suspended in a fluid, enabling easy handling and manipulation within microfluidic systems. The process begins with the loading of single cells into the nanovials for cell encapsulation. Once loaded, the cells can be assayed directly within the nanovials. The functionalized surfaces can capture secreted molecules or enable reactions that produce detectable signals, such as fluorescence, which correlate with specific cellular functions. After the phenotypic assays, nanovials containing cells of interest can be sorted using FACS techniques adapted for hydrogel particles. This allows for the rapid and precise isolation of cells based on their phenotypes as detected within the nanovials. Hydrogel nanovials combine the advantages of droplet-based systems and support the survival of adherent cancer cells. However, certain phenotypes, such as cell morphology or the organization of organelles, might be difficult to visualize, and tracking individual cells over time is also challenging. Despite these limitations, hydrogel nanovials represent a powerful tool for high-throughput single-cell functional analysis and sorting.
2.7 Single-cell in situ analysis with spatial-omics technologies
In addition to single-cell selective marking and isolation techniques, spatial-omics technologies offer alternative approaches for in situ single-cell analysis, yielding valuable insights into the microenvironmental context of individual cells. In spatial transcriptomics, methods such as 10x Genomics Visium, Slide-seq, and MERFISH are prominent. 10x Genomics Visium captures spatially resolved transcriptomes by combining tissue sections with RNA sequencing on barcoded slides, whereas Slide-seq uses DNA-barcoded beads to map gene expression profiles across tissue samples [100,101]. MERFISH (Multiplexed Error-Robust Fluorescence In Situ Hybridization) utilizes sequential hybridization and imaging to detect thousands of RNA species in fixed tissues, enabling high-throughput spatial analysis [102]. Spatial proteomics technologies like Imaging Mass Cytometry (IMC), CODEX (CO-Detection by Indexing), and MIBI (Multiplexed Ion Beam Imaging) provide high-resolution protein expression maps [103-107]. IMC integrates mass spectrometry with immunohistochemistry by using metal-tagged antibodies, allowing for multiplexed protein detection. CODEX employs DNA-barcoded antibodies and cyclic immunofluorescence to achieve high-dimensional protein profiling, while MIBI uses secondary ion mass spectrometry to map proteins at subcellular resolution. For spatial metabolomics, Imaging Mass Spectrometry (IMS) and Secondary Ion Mass Spectrometry (SIMS) are key techniques [108-110]. IMS, including methods like MALDI (Matrix-Assisted Laser Desorption/Ionization) and DESI (Desorption Electrospray Ionization), ionizes metabolites directly from tissue sections to analyze their spatial distribution. SIMS uses a focused primary ion beam to sputter secondary ions from the sample surface, providing detailed metabolite maps. These technologies collectively enable the integration of transcriptomic, proteomic, and metabolomic data, offering a comprehensive understanding of cellular molecular features within their native tissue context. Spatial-omics technologies, despite limitations such as spatial resolution constraints in achieving single-cell precision, high costs, inability to integrate with functional assays, and challenges in data quality and normalization, provide their unique value for in situ analysis of complex biological microenvironments.
3. Conclusions and Perspectives
Single-cell analysis has transformed modern biological research by providing unprecedented insights into cellular behavior and heterogeneity. Unlike conventional population-based methods, this approach examines individual cells, revealing significant variations in cellular states, responses to stimuli, and molecular signatures often obscured in bulk analyses. This level of detail is crucial for understanding complex physiological and pathological processes, enabling researchers to gain profound insights into cellular function and behavior, thus driving groundbreaking discoveries in biomedical science. In cancer research, single-cell analysis is particularly essential due to the notorious heterogeneity within tumor cell populations, which exhibit diverse gene expression, phenotypes, and functional properties. This heterogeneity, influenced by genetic mutations, epigenetic alterations, and microenvironmental factors, profoundly affects tumor progression, metastasis, and treatment resistance. Deciphering this heterogeneity is essential for advancing cancer treatment, as distinct cell subpopulations within tumors respond differently to therapies, leading to resistance and disease recurrence. By characterizing and targeting specific cell subsets, clinicians can develop personalized treatment strategies with improved efficacy. Specifically, advancements in single-cell analysis have revolutionized the ability to correlate phenotypic and genotypic data at the individual cell level, a critical step in understanding the molecular mechanisms underlying cell behaviors. Cutting-edge techniques, such as robotic aspiration, laser detachment, microraft arrays, optical tweezers, optical labeling, microfluidics, hydrogel nanovials, and droplet-based systems, offer promising strategies for high-throughput single-cell phenotypic analysis and selective cell isolation for genotypic downstream characterizations. However, challenges remain in analyzing certain phenotypes and tracking individual cells over time. Additionally, the complexity and cost of initial setups, along with limited throughput in many systems, constrain the investigation of phenotype-genotype correlations.
Future advancements in deep learning are poised to significantly enhance single-cell techniques across three primary areas: (1) Automating selective cell picking using computer vision, rather than human discretion, to markedly increase the throughput and reproducibility of single-cell isolation. (2) Optimizing the control of robotic aspiration and droplet systems to improve precision and efficiency. (3) Identifying complex patterns and relationships within single-cell data that are otherwise difficult to discern, with deep learning models integrating multi-omics data to provide comprehensive insights into cellular heterogeneity and function [111-114]. Additionally, advances in robotics operating microfluidic and other platforms will significantly boost the throughput, reliability, and reproducibility of experiments [115-118]. The data generated through robotic operations can further enhance the application of data science in system optimization, normalization, and quality control of single-cell data. Although challenges remain, rapid developments in robotics and deep learning will gradually address these issues, greatly enhancing the capability of analyzing individual cells in biomedical research. This progress is particularly valuable in cancer research, where understanding cellular heterogeneity is essential for advancing precision medicine and fostering therapeutic innovation.
Declarations
Consent for Publication
Not applicable.
Availability of Data and Material
Not applicable.
Competing Interests
The authors declare that no competing interests exist.
Declaration of Generative AI in Scientific Writing
The authors utilized ChatGPT 3.5 to enhance the readability and language of this work. Following its use, the authors thoroughly reviewed and edited the content as necessary and take full responsibility for the content of the publication.
Acknowledgement
This study was generously funded by start-up support from the UPMC Hillman Cancer Center, the Women’s Cancer Research Center (WCRC) at Magee Women’s Research Institute, the Pitt CTSI Pilot project (supported by the National Institutes of Health through Grant Number UL1TR001857), and the US National Institutes of Health grants P30 CA047904, P50 CA272218, and R35 GM150509, as well as the UPMC Competitive Medical Research Fund (CMRF) awarded to Y.-C.C.

 ,
Yushu Ma
1,2
,
Yushu Ma
1,2